Coat Assembly Control by Monomeric GTPase
개요
Sar-1(GTPase), Sar1-GEF / Sec23, Sec24 (adaptor proteins) / Sec13, Sec31 (Outer coat)
각 compartment에서 적절한 coated vesicle이 형성되려면 coat assembly가 언제, 어디서 시작될지 정밀하게 제어되어야 한다.12 이 제어에서 핵심적인 역할을 하는 것이 coat-recruitment GTPase이다. Coat-recruitment GTPase는 특정 compartment의 membrane에서 활성화되어 해당 membrane에 맞는 coat protein을 불러들이는 분자 스위치로 작동한다.
이들은 Phosphatidylinositol(PI) and Phosphoinositides(PIPs)와 함께 coat assembly의 위치 특이성을 결정하는 두 가지 주요 메커니즘 중 하나이다.
- PIP가 plasma membrane과 Golgi, endosome에서 clathrin coat를 조절하는 반면
- coat-recruitment GTPase는 Golgi, ER, endosome 모두에서 COPI, COPII, retromer coat assembly를 주도적으로 조절한다.
Coat-recruitment GTPase의 종류
| GTPase | 작동 compartment | 조절하는 coat |
|---|---|---|
| Sar1 | ER membrane | COPII coat |
| ARF proteins | Golgi membrane | COPI coat, Clathrin coat |
| Endosome membrane | Retromer coat |
각각은 서로 다른 GEF에 의해 특정 compartment에서 활성화되며, 이 GEF의 위치 자체가 coat assembly의 공간적 조절을 결정한다.
분자 스위치 메커니즘: GDP/GTP Cycle
Coat-recruitment GTPase는 monomeric GTPase의 일반적인 작동 원리를 따른다. GTP-bound 상태에서 활성, GDP-bound 상태에서 비활성 상태로 전환된다.
두 가지 조절 단백질:
- GEF (Guanine nucleotide exchange factor): GDP → GTP 교환을 촉매하여 활성화
- GAP (GTPase-activating protein): GTP hydrolysis를 촉진하여 불활성화
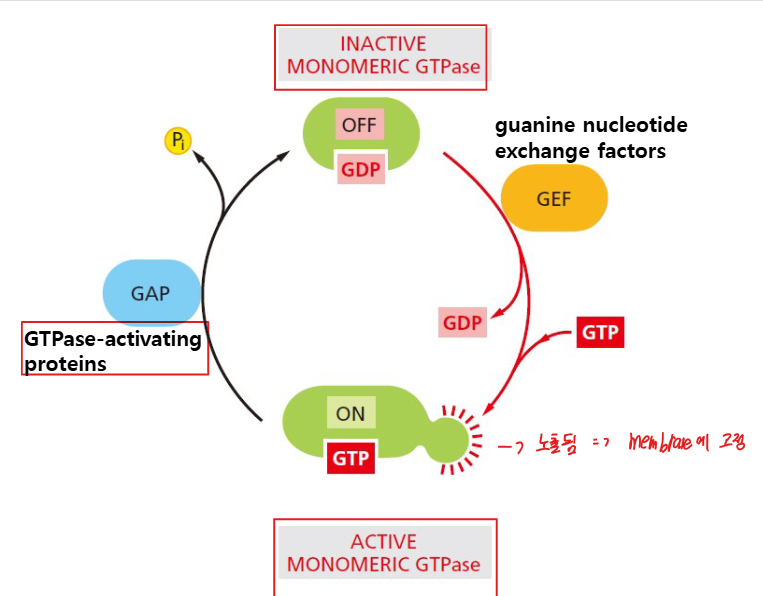 주요 특징:
주요 특징: - 세포질 내 GTP 농도가 GDP 농도보다 훨씬 높기 때문에, GDP가 방출되면 즉시 GTP가 결합됨
- 따라서 GEF가 GDP를 방출시키는 것만으로도 활성화가 일어남
- GAP는 GTPase의 가수분해 기능을 활성화 → GTPase가 자기자신에게 붙은 GTP를 분해.→GTPase의 lipid anchor 기능 불활성화
핵심 예시: Sar1에 의한 COPII Coat Assembly
COPII coat의 형성 과정은 coat-recruitment GTPase의 작동 방식을 잘 보여준다.
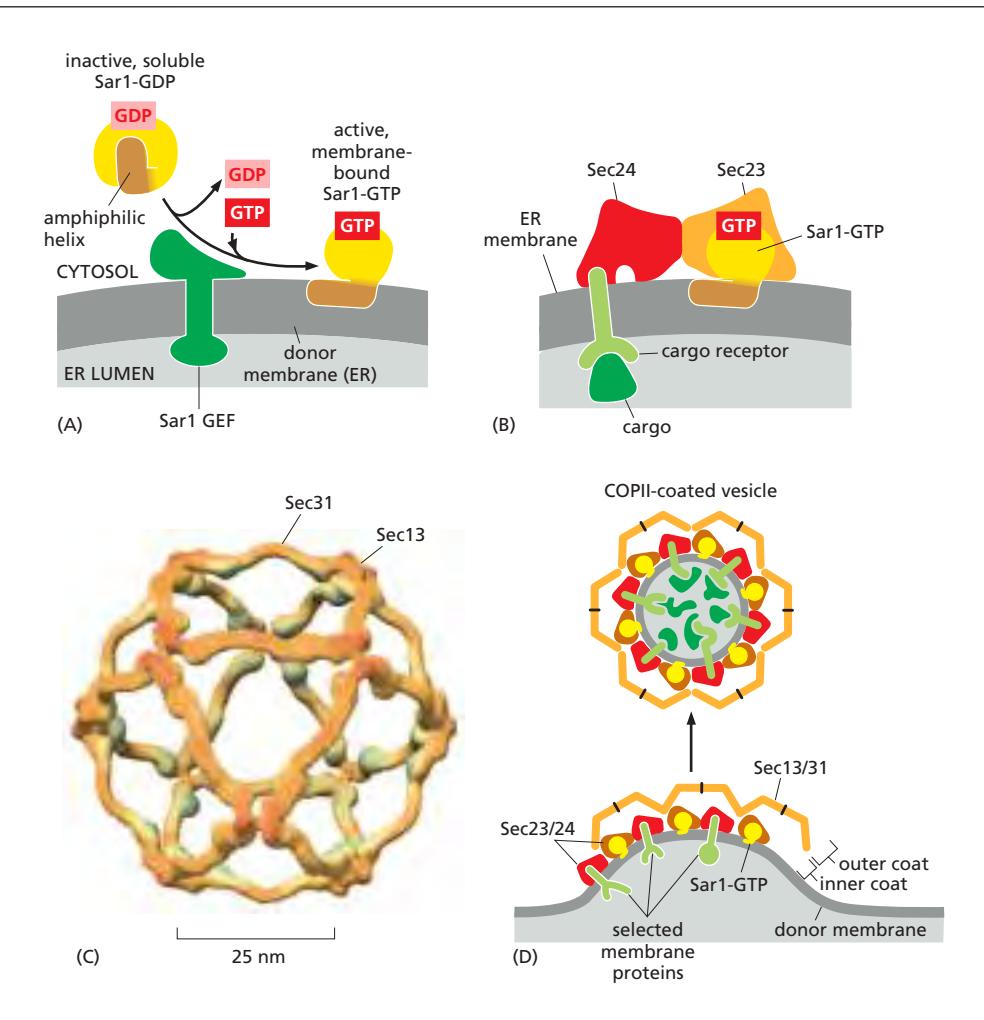 Figure 13-15: COPII-coated vesicle의 형성. (A) Sar1 GEF가 ER membrane에서 Sar1-GDP를 활성화시킨다. (B) Sar1-GTP가 inner coat인 Sec23/Sec24 복합체를 recruitment한다. (C) Outer coat인 Sec13/Sec31 복합체가 Sec23/Sec24 위에 조립된다. (D) 전체 과정의 모식도.
Figure 13-15: COPII-coated vesicle의 형성. (A) Sar1 GEF가 ER membrane에서 Sar1-GDP를 활성화시킨다. (B) Sar1-GTP가 inner coat인 Sec23/Sec24 복합체를 recruitment한다. (C) Outer coat인 Sec13/Sec31 복합체가 Sec23/Sec24 위에 조립된다. (D) 전체 과정의 모식도.
단계별 과정
1단계: Sar1 활성화
- Sar1은 평소 세포질에 Sar1-GDP 상태로 존재2
- ER membrane에 embedded된 Sar1 GEF(Sec12, 이건 Sar1이 아님.)가 세포질의 Sar1-GDP와 결합2
- GEF가 GDP를 방출시키고, 세포질에 풍부한 GTP가 즉시 결합12
- Sar1-GTP 형성12
2단계: ER membrane에 anchor
- GTP 결합으로 Sar1의 conformational change
- N-terminal의 amphiphilic(양친매성) helix가 노출2
- 이 helix가 ER membrane의 cytoplasmic leaflet에 wedge처럼 삽입
- Sar1이 ER membrane에 단단히 결합되고, 동시에 membrane curvature 유도 시작
3단계: Inner coat 조립
- Membrane-bound Sar1-GTP가 COPII adaptor coat protein인 Sec23/Sec24 복합체 모집
- Sec24는 ER exit signal을 가진 cargo receptor의 cytosolic tail에 대한 여러 binding site 보유
- Sec23/Sec24 전체 복합체의 membrane-접촉면은 완만하게 굽은 형태로, COPII vesicle의 곡률과 일치
4단계: Outer coat 조립
- Sec23/Sec24 위에 outer coat protein인 Sec13/Sec31 복합체 모집
- Sec13/Sec31은 독립적으로 삼각형, 사각형, 오각형의 cage를 형성 가능
- 적절한 크기의 vesicle을 감쌀 수 있는 symmetric cage 형성
5단계: Budding 완성
- Coat assembly가 membrane curvature를 증폭
- Vesicle neck에 dynamin 유사 machinery가 작용하여 pinch off
- COPII-coated vesicle 완성
ARF Protein에 의한 COPI Coat 및 Clathrin Coat Assembly
ARF protein (ADP-ribosylation factor)은 Golgi membrane에서 두 종류의 coat를 조절한다.
- COPI coat: Golgi cisternae 사이 retrograde transport 및 ER retrieval
- Clathrin coat (Golgi): TGN에서 endosome 및 secretory vesicle로의 transport
Sar1과 마찬가지로 ARF protein도:
- 평소 cytosol에 ARF-GDP 상태로 존재
- Golgi membrane의 ARF GEF에 의해 활성화
- GTP 결합 후 lipid anchor가 Golgi membrane에 삽입
- 해당 coat protein을 모집
GEF의 위치가 곧 coat assembly의 위치: ER의 Sar1 GEF는 COPII를 ER에서만, Golgi의 ARF GEF는 COPI/Clathrin을 Golgi에서만 활성화시킨다. 각기 다른 compartment에 위치한 GEF가 coat assembly의 공간적 특이성을 보장한다.
Coat Disassembly의 제어
Vesicle이 donor membrane에서 떨어지고 나면 coat를 빠르게 제거해야 target membrane과 융합이 가능하다. Coat disassembly 역시 coat-recruitment GTPase에 의해 시작된다.
GTP 가수분해가 trigger
GTP 가수분해가 일어나면 GTPase의 conformational change로 인해:
- 막에 삽입된 hydrophobic tail/helix가 membrane에서 빠져나옴
- GTPase가 membrane에서 분리됨
- GTPase의 이탈로 coat protein들의 결합이 불안정해짐
- Coat disassembly 시작
따라서 GTP 가수분해 속도 = coat가 유지되는 시간
Coat별 Disassembly Timing 비교
COPII coat:
- COPII coat 자체가 Sar1의 GTP hydrolysis를 촉진
- 그러나 coat가 온전히 형성되면 cargo receptor 결합 등 협동적 상호작용이 coat를 안정화
- 완전한 vesicle이 형성되는 속도가 GTP hydrolysis에 의한 disassembly보다 빠를 때만 vesicle 완성
- Vesicle pinch off 후, kinase가 coat protein을 인산화 → disassembly 완성
COPI coat:
- Vesicle membrane의 curvature 자체가 trigger
- Coat에 함께 모집된 ARF GAP가 membrane의 lipid packing density 감지
- Membrane curvature가 transport vesicle 수준에 달하면 ARF GAP 활성화
- ARF가 GTP를 가수분해 → ARF가 membrane에서 분리 → coat 즉시 disassemble
- Vesicle pinch off 직후 coat 제거
Clathrin coat:
- Coat에 co-packaged된 auxilin이 hsp70 chaperone(ATPase) 활성화
- hsp70이 ATP hydrolysis 에너지로 clathrin coat를 물리적으로 제거
- Vesicle내 phosphoinositide phosphatase가 coat adaptor를 membrane에서 이탈시킴
Timing의 중요성
Coat는 vesicle이 완전히 형성되기 전에 너무 일찍 제거되어서는 안 된다. 그렇지 않으면 반쯤 형성된 bud가 다시 donor membrane으로 흡수된다. 이에 대한 해결책으로:
- Coat-recruitment GTPase가 coat assembly를 시작하는 동시에 disassembly timer를 세팅
- Cargo receptor와의 결합, 협동적 coat protein 상호작용이 GTP hydrolysis보다 coat를 안정화
- 완전한 vesicle이 형성될 때까지 coat가 유지됨
요약
Coat-recruitment GTPase는 특정 compartment membrane에 위치한 GEF에 의해 그 자리에서만 활성화되어, 해당 membrane에 적합한 coat protein을 불러들이는 분자 스위치다. Sar1은 ER에서 COPII coat를, ARF protein은 Golgi에서 COPI 및 clathrin coat를, Rab7은 endosome에서 retromer coat를 조절한다. 활성화된 GTPase는 amphiphilic helix나 lipid anchor를 통해 membrane에 삽입되고, 이후 coat protein의 단계적 조립을 유도한다. GTP 가수분해는 coat disassembly의 trigger로 작용하며, 그 속도가 vesicle이 유지되는 시간을 결정한다.
Footnotes
-
2022 중간 24번 — Sar1의 GEF에 의한 활성화, amphiphilic helix의 ER membrane 삽입, Sec23/24→Sec13/31 순서의 COPII coat 조립이 정답 근거로 활용됨. ↩ ↩2 ↩3
-
2023 중간 18번 — ①번 선지: A는 항상 soluble 단백질로 존재한다 (틀림, GAP(A)는 일부 막 결합 형태로도 기능); ②번 선지: B는 비활성화 G 단백질을 활성화시킨다 (틀림, GEF(B)는 Sar1-GDP에서 GDP를 방출시켜 GTP가 결합되게 하는 것으로 활성화, 설명은 맞지만 ‘비활성화’의 대상이 Sar1-GDP이므로 맞음 — 그러나 실제 해설에서는 이것이 맞는 선지로 나옴); ③번 선지: B는 pinch off 과정에도 중요한 역할을 할 수 있다 (맞음, GEF(B=Sec12)는 Sar1을 활성화하고 Sar1-GTP의 amphipathic helix가 ER membrane 삽입 → coat assembly → pinch off에 기여, 정답); ④번 선지: Sar1은 C의 소수성 그룹으로 이루어진 막을 통해 막에 결합할 수 있다 (틀림, Sar1은 amphiphilic(양친매성) helix로 결합, farnesyl 같은 소수성 그룹이 아님); ⑤번 선지: A 단백질은 모든 G 단백질에 대해 보편적으로 작용하는 하나의 단백질이다 (틀림, GAP는 특정 G 단백질에 특이적으로 작용). ↩ ↩2 ↩3 ↩4 ↩5 ↩6