세포골격 (1) 정리 — Actin Filaments & Skeletal Muscle
7주차 강의 (Cytoskeleton 1) 전체 내용 요약. 상세 내용은 100_Actin filaments_MOC, 400_Skeletal Muscle Cell_MOC 참조.
Function and Dynamics of the Cytoskeleton
세포골격(cytoskeleton)은 세포 내부를 구조적으로 조직하는 단백질 필라멘트 네트워크이다. 주요 기능은 다음과 같다.
- Cell shape and support — 세포 형태 유지
- Cell movement — 세포 이동 및 내부 운동
- Intracellular transport of vesicles and organelles — 세포 내 수송 경로 제공
- Keep organelles in place — 세포소기관 위치 고정
- Assist in cell signaling — 세포 신호전달 보조
세포골격을 이루는 세 종류의 필라멘트는 동일한 단백질 풀을 이용하면서도 세포 상태에 따라 완전히 다른 구조로 신속히 재조직된다.
Cytoskeleton during Cell Division
→ Cytoskeleton during cell division 참조
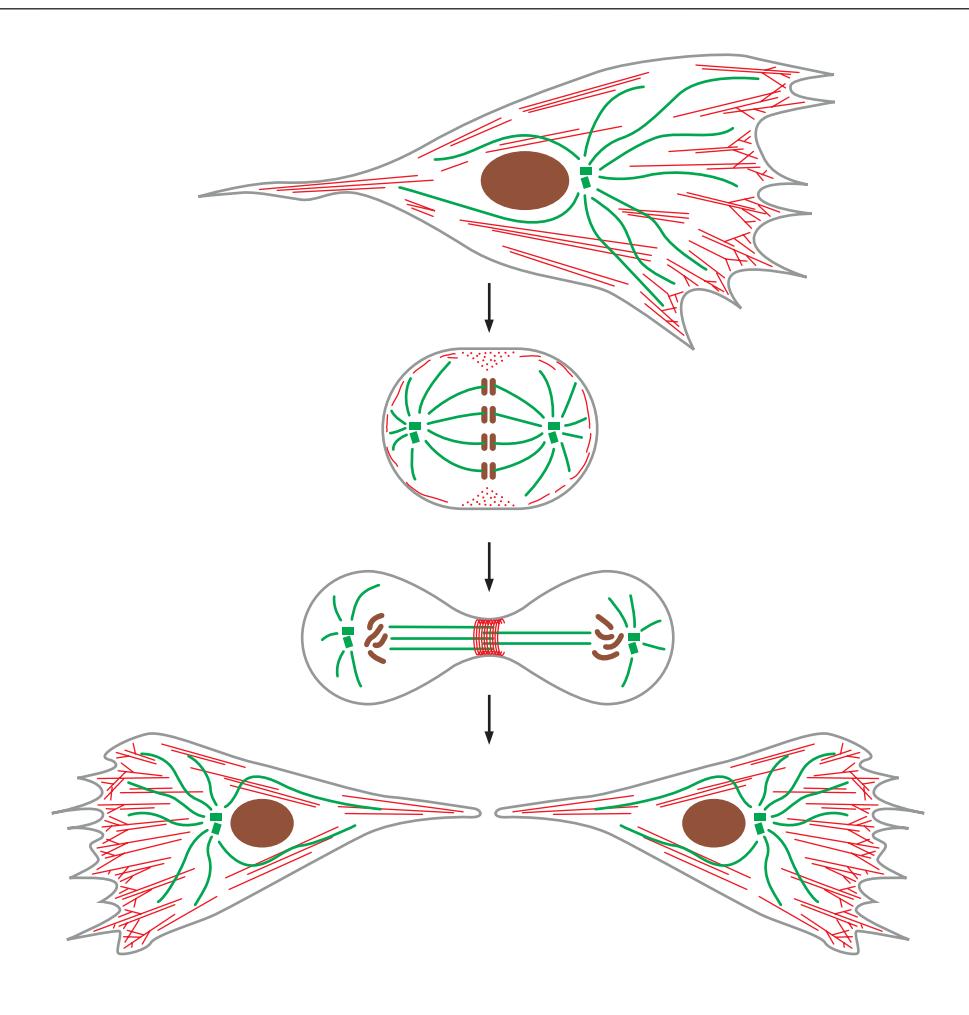 Fig 16–2. 세포 분열에 따른 세포골격 재조직. 이동 중인 fibroblast에서 분열까지.
Fig 16–2. 세포 분열에 따른 세포골격 재조직. 이동 중인 fibroblast에서 분열까지.
같은 세포골격 단백질들이 세포 주기 단계에 따라 전혀 다른 고차 구조를 형성한다.
Interphase
- Actin filament: 세포 표면 아래 얇은 cortical network, stress fiber, lamellipodium, filopodium 등 다양한 구조 형성
- Microtubule: centrosome에서 방사형으로 뻗어(aster) 세포소기관 위치 결정 및 intracellular transport 경로 제공; mother centriole이 basal body로 기능하여 primary cilia 형성
- Intermediate filament: 핵 주위 cage 및 세포질 전반에 걸쳐 기계적 강도 제공
유사분열 및 세포질분열
- Microtubule: 두 centrosome에서 mitotic spindle 형성 → 염색체를 양 극으로 분리
- Actin filament: 세포질분열 시 actin + myosin II로 구성된 contractile ring 형성 → 수축하여 세포를 두 딸세포로 분리
- Intermediate filament: 유사분열 진입 시 kinase에 의한 인산화로 부분 해체; nuclear lamin은 핵막 붕괴와 함께 해체 후 분열 후 재조립
Actin Filaments
개요
Actin filaments (microfilaments)는 actin 단백질의 나선형 중합체이다. 직경 8 nm의 유연한 구조로, linear bundle, 2차원 network, 3차원 gel 등 다양한 형태로 조직된다. 세포 전체에 분포하나 plasma membrane 바로 아래 cortex에 가장 고농도로 집중된다.
Structure of Actin Filaments
→ structures of an actin monomer and actin filament 참조
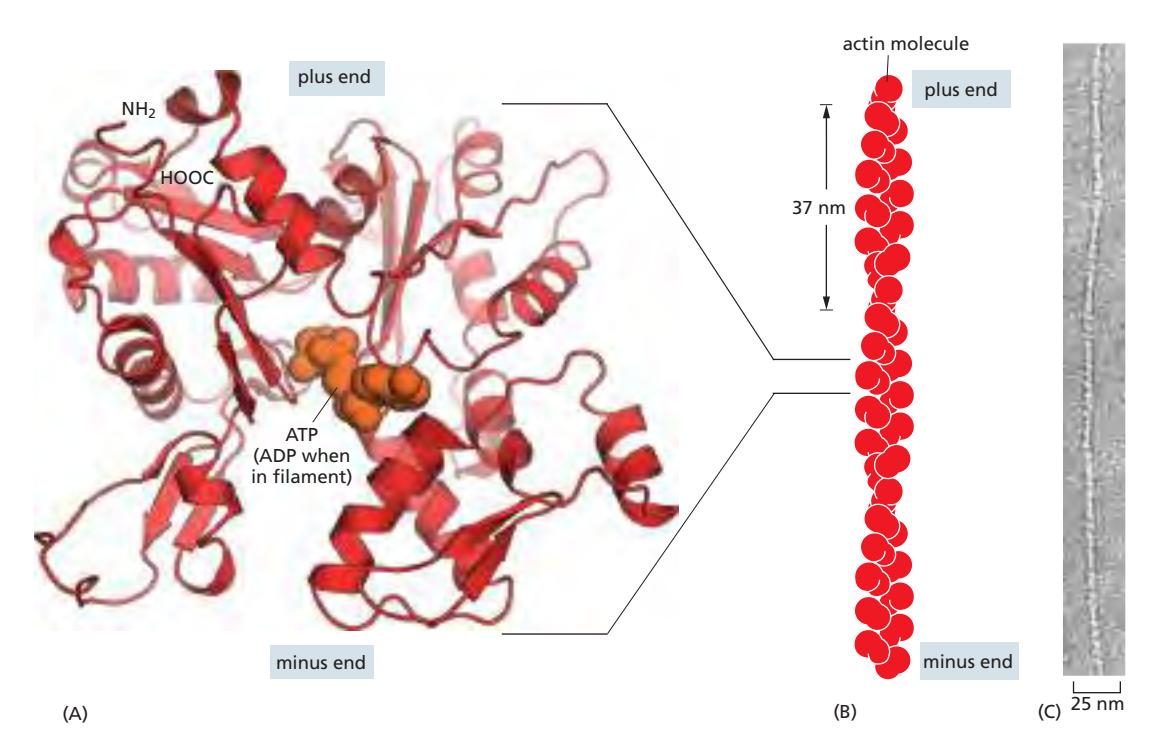 Fig 16–9. Actin 단량체와 필라멘트 구조.
Fig 16–9. Actin 단량체와 필라멘트 구조.
G-actin (globular actin)
- 약 375개의 아미노산으로 구성된 구형 단량체
- ATP 또는 ADP에 결합한 상태로 존재하며, ATP-actin은 중합되기 쉽고 ADP-actin은 탈중합되기 쉽다
F-actin (filamentous actin)
- G-actin이 중합하여 형성되는 우회전성(right-handed) 이중 나선 구조
- 직경 8 nm, 피치(pitch) 37 nm
- 극성(polarity)을 가짐:
- Plus end (barbed end): 단량체 추가가 빠른 쪽
- Minus end (pointed end): 단량체 추가가 느린 쪽
ATP Hydrolysis and Treadmilling
→ treadmilling 참조
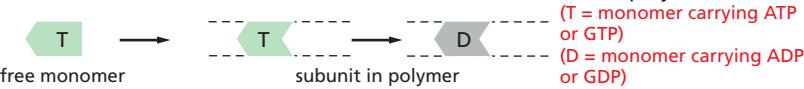
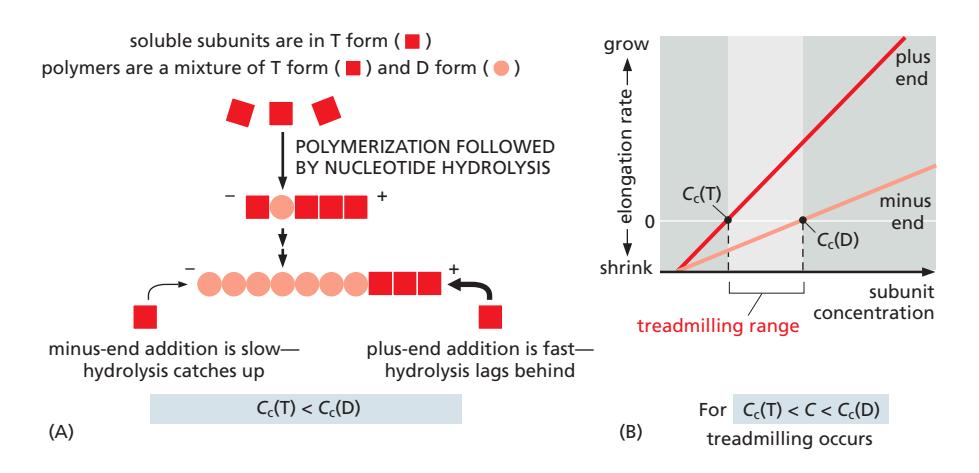 Fig 16–14. Treadmilling. (A) Plus end terminal subunit은 T형(ATP), minus end는 D형(ADP) 유지. (B) Cc(T) < C < Cc(D) 조건에서 treadmilling 발생.
Fig 16–14. Treadmilling. (A) Plus end terminal subunit은 T형(ATP), minus end는 D형(ADP) 유지. (B) Cc(T) < C < Cc(D) 조건에서 treadmilling 발생.
ATP-actin(T형)은 critical concentration Cc(T)가 낮아 중합되기 쉽고, ADP-actin(D형)은 Cc(D)가 높아 탈중합되기 쉽다.
- Plus end: 중합 속도 > 가수분해 속도 → terminal subunit이 항상 T형 유지 → 성장
- Minus end: 가수분해 속도 > 중합 속도 → terminal subunit이 D형 유지 → 탈중합
- Minus end에서도 T form actin이 결합할 수 있으나, D form으로의 가수분해 속도가 더 빨라 결과적으로 탈중합
- 자유 subunit 농도 C가 **Cc(T) < C < Cc(D)**인 조건에서 treadmilling 발생: plus end에서 조립, minus end에서 분해가 동시에 일어나 필라멘트가 앞으로 흐르는 것처럼 이동
세포 내 가속화:이는 cofilin이 ADP-actin 필라멘트에 선택적으로 결합하여 탈중합을 수십 배 가속하기 때문이다.
Chemical Inhibitors of Actin and Microtubules
→ Chemical Inhibitors of Actin and Microtubules 참조
세포골격 필라멘트를 안정화하거나 불안정화하는 화학물질들은 세포 내 역할 연구와 항암 치료에 활용된다. 이 약물들은 필라멘트가 역동적 평형(dynamic equilibrium) 상태에서 기능함을 보여준다.
| Chemical | Effect on filaments | Mechanism | Original source |
|---|---|---|---|
| Actin | |||
| Latrunculin | Depolymerizes | Binds actin subunits | Sponges |
| Cytochalasin B | Depolymerizes | Caps filament plus ends | Fungi |
| Phalloidin | Stabilizes | Binds along filaments | Amanita mushroom |
| Microtubules | |||
| Taxol (paclitaxel) | Stabilizes | Binds along filaments | Yew tree |
| Nocodazole | Depolymerizes | Binds tubulin subunits | Synthetic |
| Colchicine | Depolymerizes | Caps filament ends | Autumn crocus |
Actin 저해제:
- Latrunculin (해면동물 유래): actin subunit에 직접 결합하여 중합 방해
- Cytochalasin B (진균 유래): actin 필라멘트의 plus end를 capping하여 subunit 추가 차단 → 탈중합
- Phalloidin (Amanita 독버섯 유래): 필라멘트 측면 전체에 결합하여 탈중합 억제 → 안정화. 형광 표지 phalloidin은 세포 내 actin 필라멘트 시각화의 표준 실험 도구로 사용됨(마커)
Microtubule 저해제:
- Taxol / paclitaxel (주목나무 유래): microtubule 측면에 결합하여 안정화. 암세포의 mitosis에서 방추사 형성 후 disassembly를 방해하여 염색체를 양 극으로 당기지 못하게 함 → 항암 효과. 특히 유방암·난소암 치료에 널리 사용됨
- Nocodazole (합성): tubulin subunit에 결합하여 중합 억제 → 탈중합
- Colchicine (가을 크로커스 유래): tubulin subunit에 결합하여 필라멘트 말단을 cap → 중합 저해
중합 저해제(nocodazole, colchicine)와 탈중합 저해제(taxol) 모두 mitotic spindle의 올바른 기능에 microtubule dynamics가 필수적이기 때문에 분열 세포를 우선적으로 사멸시킨다.
Actin Arrays in a Cell
→ actin arrays 참조
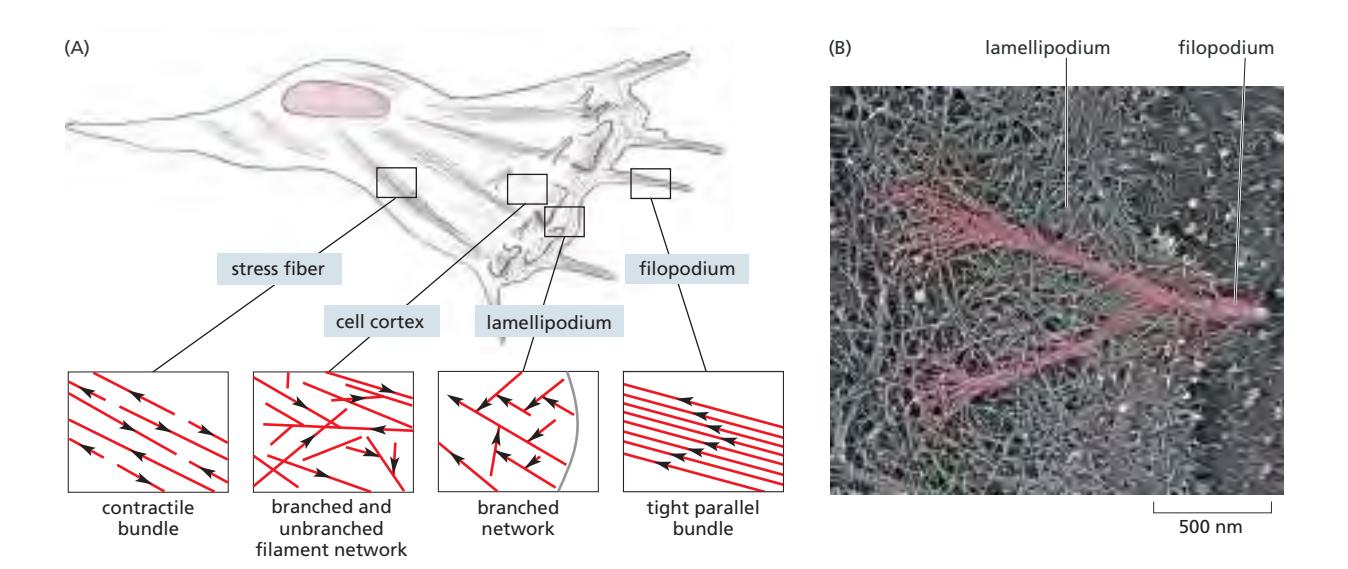 Fig 16–21. 이동 중인 fibroblast의 actin array. 화살표가 향하는 방향이 minus end 쪽.
Fig 16–21. 이동 중인 fibroblast의 actin array. 화살표가 향하는 방향이 minus end 쪽.
세포 내 actin 필라멘트는 다양한 accessory protein에 의해 네 가지 주요 array로 조직되며, 각각 Rho GTPase family에 의해 조절된다.
| Array 유형 | 구조적 특징 | 기능 | 조절 GTPase |
|---|---|---|---|
| Dendritic network (lamellipodium) | Arp2/3가 기존 filament에서 70° 각도로 새 filament nucleation; sheet-like | 세포 가장자리 앞쪽 돌출; 방향성 있음 | Rac |
| Tight parallel bundle (filopodium) | Formin이 unbranched straight filament 형성; spike형 | 환경 탐색, 신경 성장원뿔 유도; plus end가 세포 끝을 향함 | Cdc42 |
| Contractile bundle (stress fiber) | 반대 극성의 actin + myosin II; bundle 형태; 양방향 | focal adhesion에 연결되어 기질에 traction 생성 | Rho |
| Gel-like network (cell cortex) | plasma membrane 아래 2D·3D network | 세포에 기계적 강도 제공, membrane 형태 유지; 명확한 방향성 없음 | — |
Rho GTPase Family
→ Rho family 참조
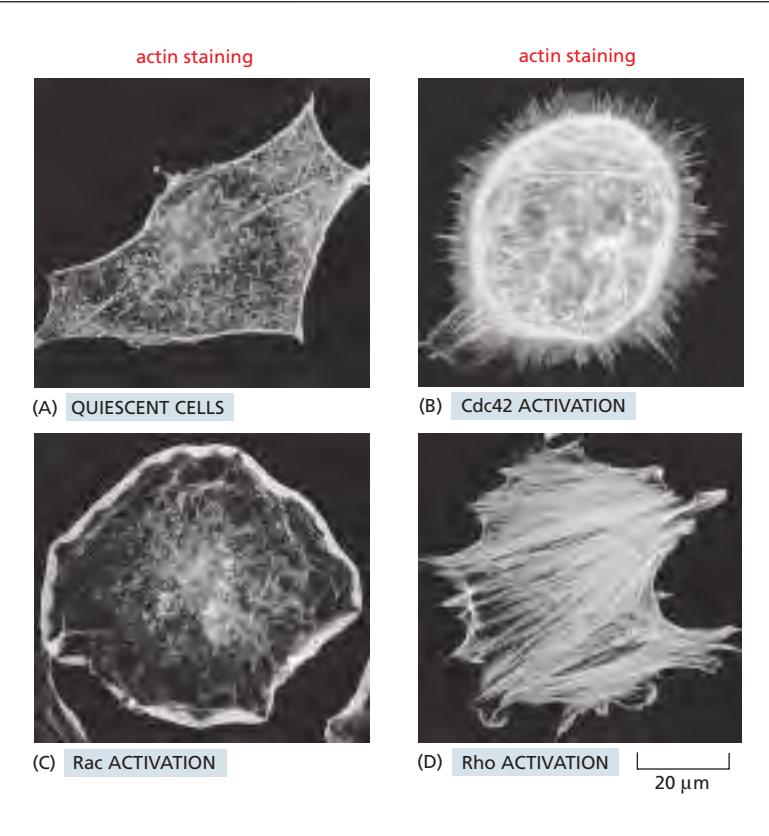 Fig 16–84. Cdc42, Rac, Rho의 활성화 효과. (A) Quiescent state(serum-starved). (B) 활성형 Cdc42 → filopodia. (C) 활성형 Rac → 전 둘레에 lamellipodium. (D) 활성형 Rho → stress fiber.
Fig 16–84. Cdc42, Rac, Rho의 활성화 효과. (A) Quiescent state(serum-starved). (B) 활성형 Cdc42 → filopodia. (C) 활성형 Rac → 전 둘레에 lamellipodium. (D) 활성형 Rho → stress fiber.
Rho family GTPase는 세포 극성 확립과 actin cytoskeleton 재구성의 핵심 신호전달 분자이다.
분자 스위치 메커니즘: GTP-결합 활성형 ↔ GDP-결합 비활성형 전환
- GEF (Guanine nucleotide Exchange Factor): GDP → GTP 교환 → 활성화
- GAP (GTPase Activating Protein): GTP 가수분해 촉진 → 불활성화
- GDI (Guanine nucleotide Dissociation Inhibitor): GDP-결합형 고정 → GEF 작용 억제
세 주요 구성원:
| GTPase | 유도 actin 구조 | 핵심 effector |
|---|---|---|
| Cdc42 | Filopodia | |
| Rac | Lamellipodia | |
| Rho | Stress fiber + Focal adhesion | |
Cell Migration by Actin Cytoskeleton
→ cell migration by actin cytoskeleton 참조
Mesenchymal cell migration (lamellipodium 기반):
- Protrusion — actin 중합 → plasma membrane 앞으로 밀어냄(lamellipodium 형성)
- Attachment — 새 focal adhesion 형성 → 기질에 부착
- Contraction — 뒤쪽 actin-myosin 수축 → 세포 몸통을 앞으로 당김
- Detachment — 뒤쪽 adhesion 해체 → 앞으로 이동
Protrusion of the Actin Meshwork at the Leading Edge:
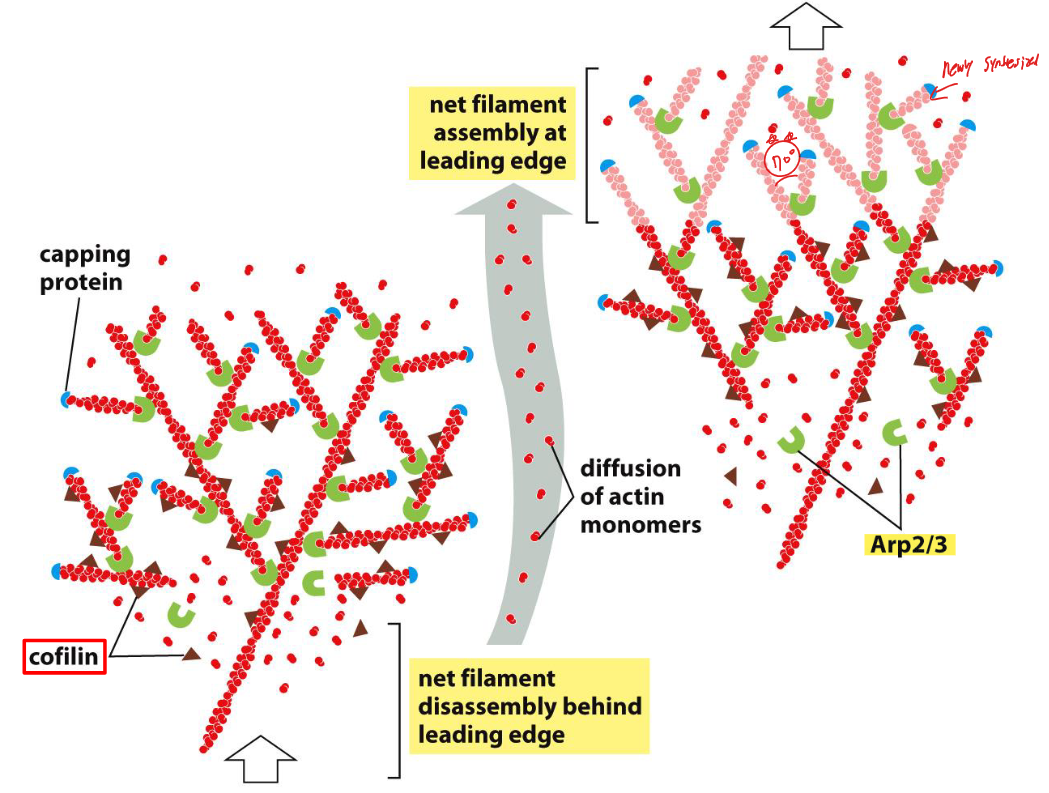 Fig 16–80. Leading edge에서 actin meshwork protrusion 모델.
Fig 16–80. Leading edge에서 actin meshwork protrusion 모델.
Leading edge에서의 actin meshwork protrusion은 다음 단계로 이루어진다:
- Nucleation: Arp2/3 complex가 leading edge 전방에서 기존 필라멘트 측면에 새 필라멘트를 nucleate (주로 70° 각도로 분기)
- Elongation: 새 필라멘트의 plus end가 길어지면서 plasma membrane을 앞으로 밀어냄. Network 뒤쪽의 anchorage 덕분에 가능
- Capping: 일정한 속도로 capping protein이 plus end를 막음
- Depolymerization: 필라멘트 내 ATP → ADP 가수분해 후 cofilin이 ADP-actin 필라멘트를 선택적으로 인식하여 탈중합 가속
결과: front에서는 net 조립, rear에서는 net 분해가 공간적으로 분리 → actin 필라멘트 network 전체는 앞으로 전진하나, 개별 필라멘트는 기질에 대해 정지 상태 유지
Major Accessory Proteins of the Actin Cytoskeleton
→ major accessory proteins of the actin cytoskeleton 참조
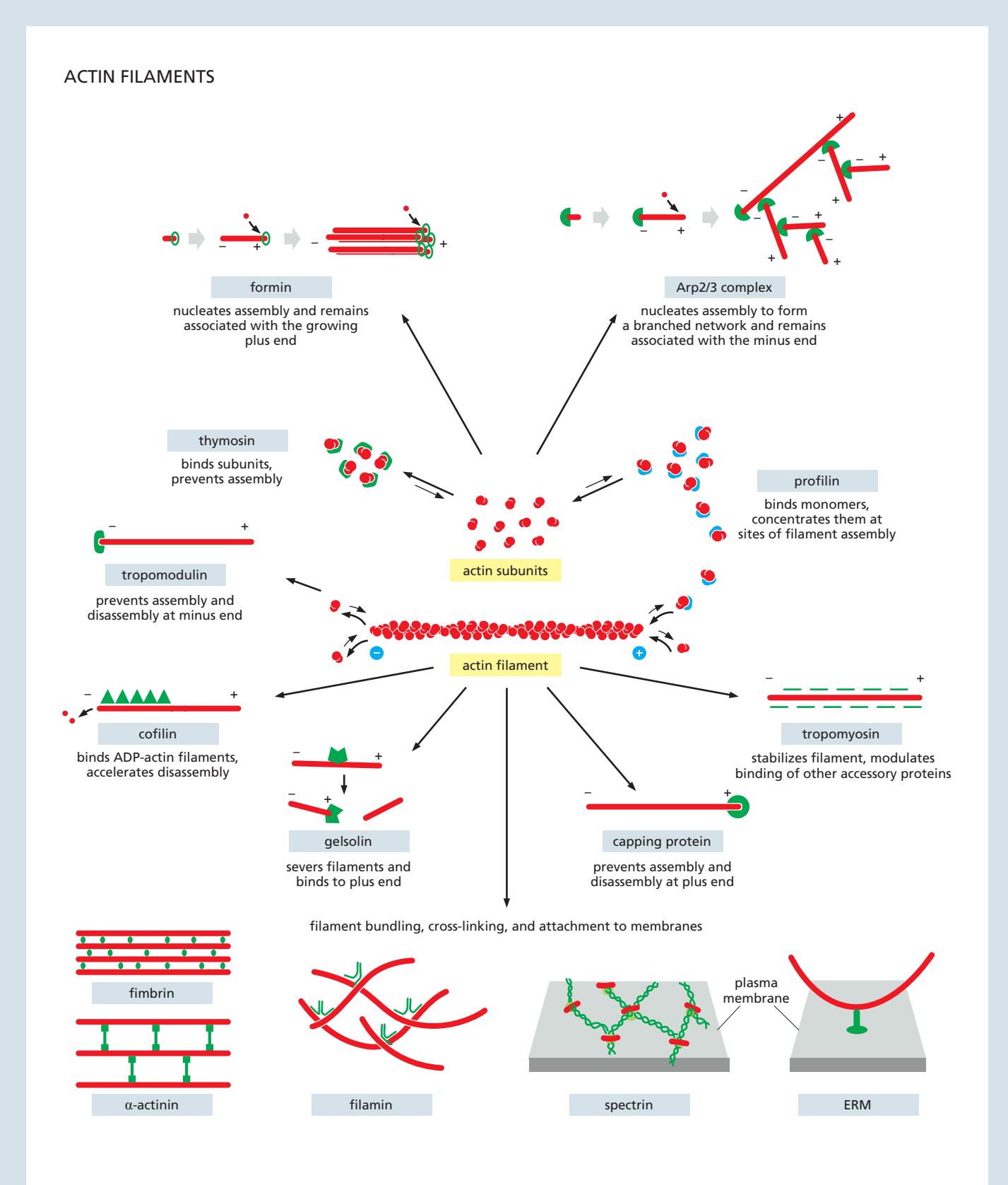 Fig 16–11. Actin cytoskeleton 주요 accessory protein 종류.
Fig 16–11. Actin cytoskeleton 주요 accessory protein 종류.
대부분의 세포에는 100개 이상의 actin-binding protein이 존재한다. 기능별 분류:
Nucleating Proteins
- Arp2/3 complex: 기존 필라멘트 측면에서 70° 각도로 새 필라멘트 분기(treelike branched network 형성). Minus end에 결합하여 남고 plus end에서 빠른 elongation. Nucleation-promoting factor (NPF) 활성화 필요
Filament End 조절 단백질
- Capping protein: actin 필라멘트 plus end를 막아 elongation 방지. Lamellipodium에서 일정 속도로 plus end를 막아 필라멘트 길이 제한 및 분기 network 구조 유지
- Tropomodulin: minus end capping 단백질. 근육 sarcomere의 thin filament에서 특히 중요
Severing / Depolymerization 단백질
- Cofilin: . 오래된(ADP-함유) 필라멘트를 선택적으로 분해하여 leading edge에서 방향성 있는 actin network 성장 가능하게 함
 Fig 16–16. Cofilin에 의한 actin 필라멘트 비틀림. Cofilin 코팅 필라멘트(B)가 순수 actin 필라멘트(A)보다 더 단단히 꼬임.
Fig 16–16. Cofilin에 의한 actin 필라멘트 비틀림. Cofilin 코팅 필라멘트(B)가 순수 actin 필라멘트(A)보다 더 단단히 꼬임.
Cross-linking / Bundling 단백질
- Spectrin: cortical cytoskeleton 구성 단백질, tetramer 형태
Stabilizing 단백질
- Tropomyosin: actin 필라멘트 홈을 따라 길게 결합하여 cofilin으로부터 보호, 특정 구조에 기능 부여. 근육 수축 조절에서 troponin과 함께 작용; Ca²⁺ 낮을 때 myosin head의 actin filament 결합부위를 가림
- Nebulin: actin filament 측면에 나선형으로 부착, sarcomere에서 actin의 일정한 길이 유지
Myosin Superfamily
→ Myosin II 참조
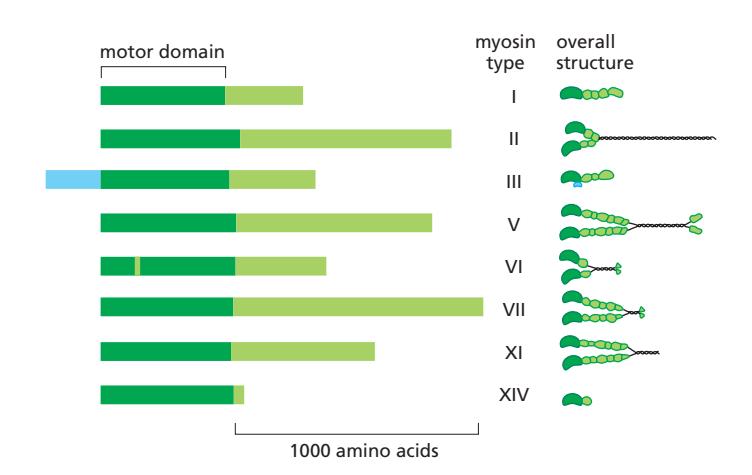 Fig 16–35. Myosin superfamily. 모든 myosin은 공통 motor domain(진한 녹색)을 공유하나 tail domain은 매우 다양하다.
Fig 16–35. Myosin superfamily. 모든 myosin은 공통 motor domain(진한 녹색)을 공유하나 tail domain은 매우 다양하다.
Myosin 계열은 최소 37개의 distinct family로 이루어진 대규모 superfamily이다. 모든 myosin heavy chain은 N-terminus에 공통적인 myosin motor domain을 가진다.
- Myosin-I (one-headed): microvilli 같은 actin 구조 돌출과 endocytosis에 관여
- Myosin-II (two-headed): 근육 수축의 핵심; 비근육 세포에서도 짧은 bipolar filament 형성
- Myosin-V: processively하게 actin 필라멘트를 따라 이동, organelle 수송
- Myosin-VI: minus end 방향으로 이동하는 유일한 myosin
Myosin II Structure
→ Myosin II 참조
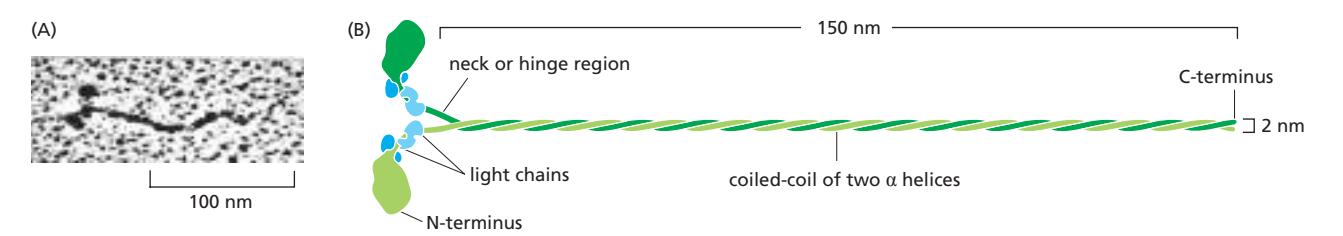 Fig 16–21. Myosin II 분자 구조. Head domain, neck(lever arm), coiled-coil tail.
Fig 16–21. Myosin II 분자 구조. Head domain, neck(lever arm), coiled-coil tail.
구조:
- Myosin II는 두 개의 heavy chain과 두 쌍의 light chain으로 구성된 elongated 단백질이다
- 각 heavy chain은 N-terminus에 globular head domain을 가지며, 그 뒤로 긴 α-helical 서열이 extended coiled-coil을 형성하여 heavy chain dimerization을 매개한다 (150 nm × 2 nm)
- 두 light chain은 N-terminal head domain 근처에 결합
- 긴 coiled-coil tail은 다른 myosin 분자의 tail과 결합
Bipolar Thick Filament:
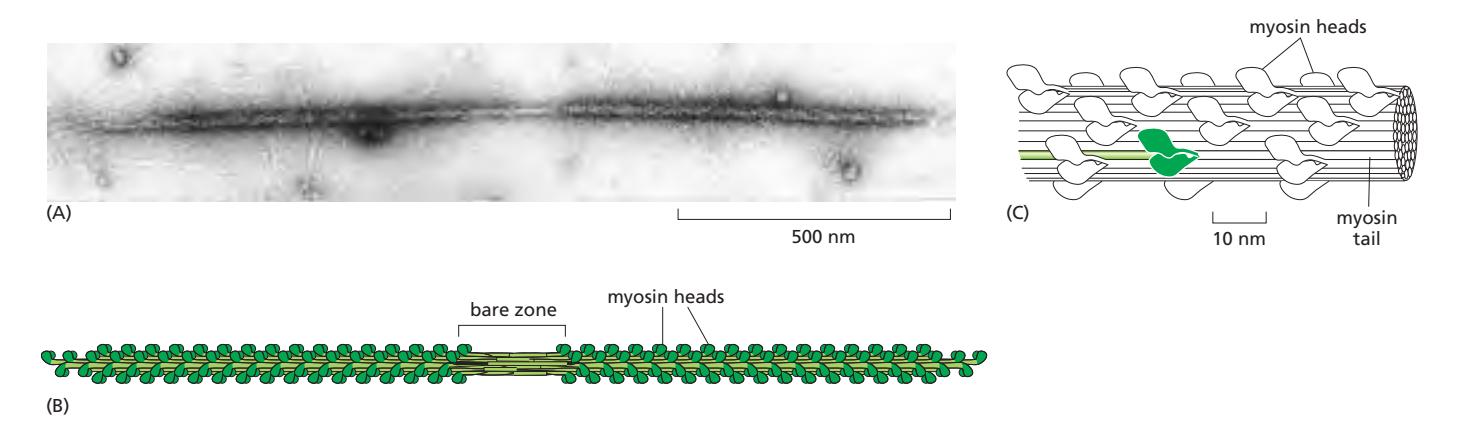 Fig 16–22. Myosin II bipolar thick filament. 중앙에 myosin tail만으로 이루어진 bare zone이 보인다.
Fig 16–22. Myosin II bipolar thick filament. 중앙에 myosin tail만으로 이루어진 bare zone이 보인다.
- 골격근에서 myosin II 분자들은 tail-tail 상호작용으로 큰 bipolar thick filament 형성
- 두꺼운 필라멘트 양 끝에 수백 개의 myosin head가 반대 방향으로 배치
- 중앙에는 myosin tail만으로 이루어진 bare zone 존재
- 이 반대 방향 배치 덕분에 두 방향의 actin 필라멘트를 서로 향해 잡아당겨 근육 단축 가능
- Myosin head는 사선으로 연결되어 텐션 유지
Myosin II Cycle
→ Myosin II cycle 참조
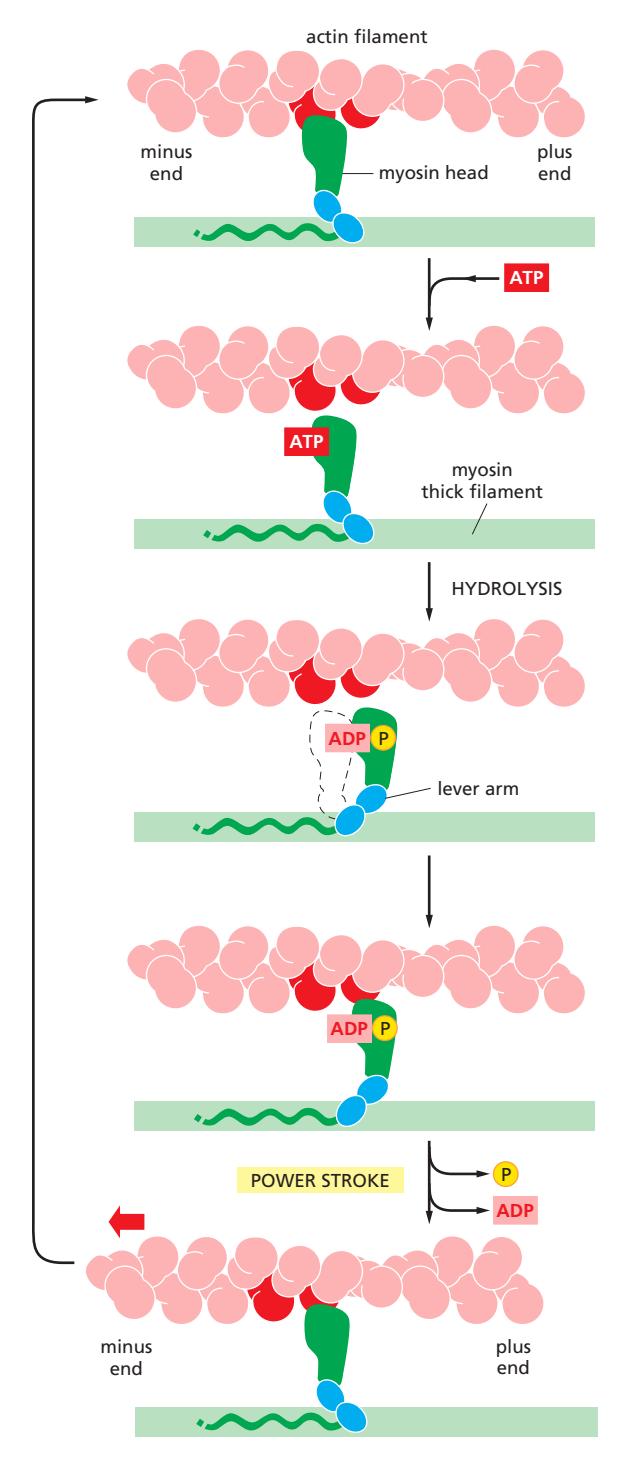 Fig 16–24. Myosin II mechanochemical cycle.
Fig 16–24. Myosin II mechanochemical cycle.
Myosin II는 ATP 가수분해를 역학적 일로 전환하기 위해 actin 필라멘트와 반복적 상호작용 주기를 거친다. 각 thick filament의 myosin head들은 ATP 주기 시간의 5%만 actin에 결합하여 힘을 발생시키므로, 많은 head들이 빠른 순서로 같은 thin filament에 작용할 수 있다. Myosin head는 minus end에서 plus end 방향으로 이동하며, actin 필라멘트를 약 5 nm 이동시킨다.
주기 5단계:
- Attached (Rigor state): Myosin head가 ADP 없이 actin에 강하게 결합된 초기 상태. Rigor mortis(사후경직)의 근육 상태
- Released: ATP가 myosin head에 결합 → head의 actin에 대한 친화력 급격히 감소 → actin에서 분리
- Cocked: ATP → ADP + Pi 가수분해. ADP와 Pi는 head에 고정 유지. Lever arm이 “cock”되어 고에너지 구조로 전환 → actin 필라멘트의 새 위치에 결합 준비
- Force-Generating (Power stroke): Myosin head가 actin 필라멘트의 새 위치에 결합 → Pi 방출 → lever arm이 앞에서 뒤로 흔들리는 power stroke → actin 필라멘트 약 5 nm 이동
- Attached (Rigor state): ADP 방출 후 rigor state로 복귀
Skeletal Muscle
Skeletal Muscle Cells
골격근 세포(muscle fiber)는 다수의 myoblast가 융합하여 형성된 거대 다핵 세포이다. 성인 인간에서 직경 약 50 μm, 길이 최대 수 cm에 달한다. 핵은 세포 주변부에 위치한다.
Myofibrils and Sarcomere
→ myofibrils 참조
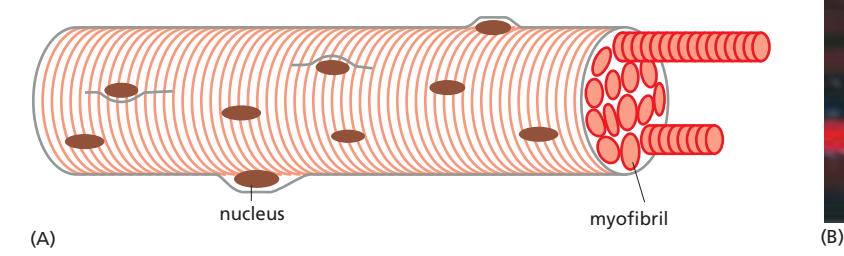 Fig 16–28. 골격근 세포 구조. 근섬유는 많은 myofibril로 구성되고, 각 myofibril은 직렬로 배열된 sarcomere로 이루어진다.
Fig 16–28. 골격근 세포 구조. 근섬유는 많은 myofibril로 구성되고, 각 myofibril은 직렬로 배열된 sarcomere로 이루어진다.
 Insect flight muscle cross section. Actin과 myosin filament가 정렬되어 있음. 커다란 단면 하나가 1 myofibril (직경 1~2 μm).
Insect flight muscle cross section. Actin과 myosin filament가 정렬되어 있음. 커다란 단면 하나가 1 myofibril (직경 1~2 μm).
골격근 세포는 myofibril이라 불리는 긴 수축성 구조물들로 채워져 있다. 각 myofibril은 수천 개의 sarcomere가 직렬로 연결된 구조이며, sarcomere는 actin thin filament와 myosin thick filament가 고도로 정렬된 최소 수축 단위이다 (길이: 2 μm).

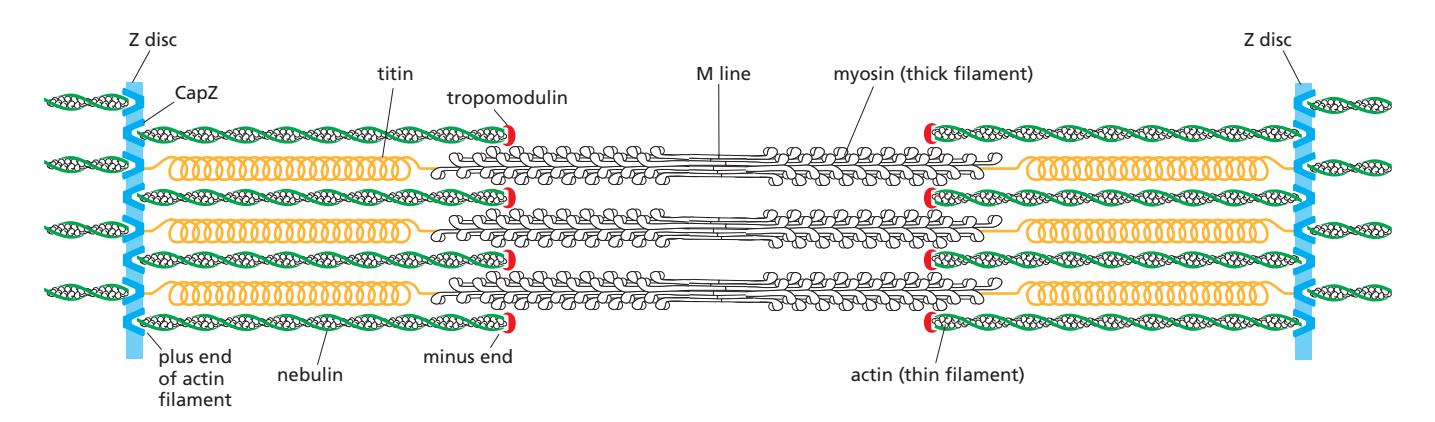
Sarcomere의 밴드 구조 (전자현미경 줄무늬):
| 구조 | 특징 |
|---|---|
| Z disc (Z line) | Sarcomere 양 경계; α-actinin 포함; actin plus end(barbed end) 고정 |
| I band (Light band) | Z disc 양쪽; actin thin filament만 포함 → 밝게 보임 |
| A band (Dark band) | Myosin thick filament 전 구간 → 어둡게 보임; actin 겹치는 구간 포함 |
| H zone | A band 중앙; myosin만 있고 actin이 겹치지 않는 구간 |
| M line | Thick filament 중앙; myosin tail 연결 단백질로 구성 |
Sliding Filament Model: 근육 수축 시 sarcomere 길이가 줄어든다. Actin thin filament와 myosin thick filament 자체의 길이 변화 없이, 두 필라멘트가 서로를 향해 미끄러진다. Myosin head의 반복적 power stroke가 actin 필라멘트를 M line 쪽으로 잡아당겨 Z disc 간 거리가 좁아진다.
Accessory Proteins in Sarcomere
→ Organization of accessory proteins in a sarcomere 참조
Sarcomere에는 actin과 myosin II 외에 다수의 accessory protein이 정밀하게 배치된다:
| 단백질 | 위치 / 기능 |
|---|---|
| Titin | Z disc ~ M line; myosin thick filament와 연결; 탄성 있어 sarcomere 수축·이완 시 길이 변화 → 분자 스프링 역할, thick filament를 정중앙에 위치, 과도한 stretching 제한 |
| Nebulin | Thin filament 전장과 정확히 일치하는 길이; thin filament 측면에 배치 → 분자 자(ruler) 역할, actin 길이 규격화 |
| Tropomodulin | Thin filament minus end capping → 필라멘트 길이 안정화 |
| CapZ | Thin filament plus end를 Z disc에 고정 |
| α-Actinin | Z disc에 존재; actin 필라멘트 간 cross-linking |
| Tropomyosin | Actin 필라멘트 홈을 따라 결합; troponin과 함께 Ca²⁺ 신호에 따른 수축 조절 |
| Troponin | Ca²⁺ 감지 복합체; tropomyosin 위치 변경으로 myosin 결합 부위 노출 조절 |
T Tubules and Sarcoplasmic Reticulum
→ T tubules, sarcoplasmic reticulum 참조
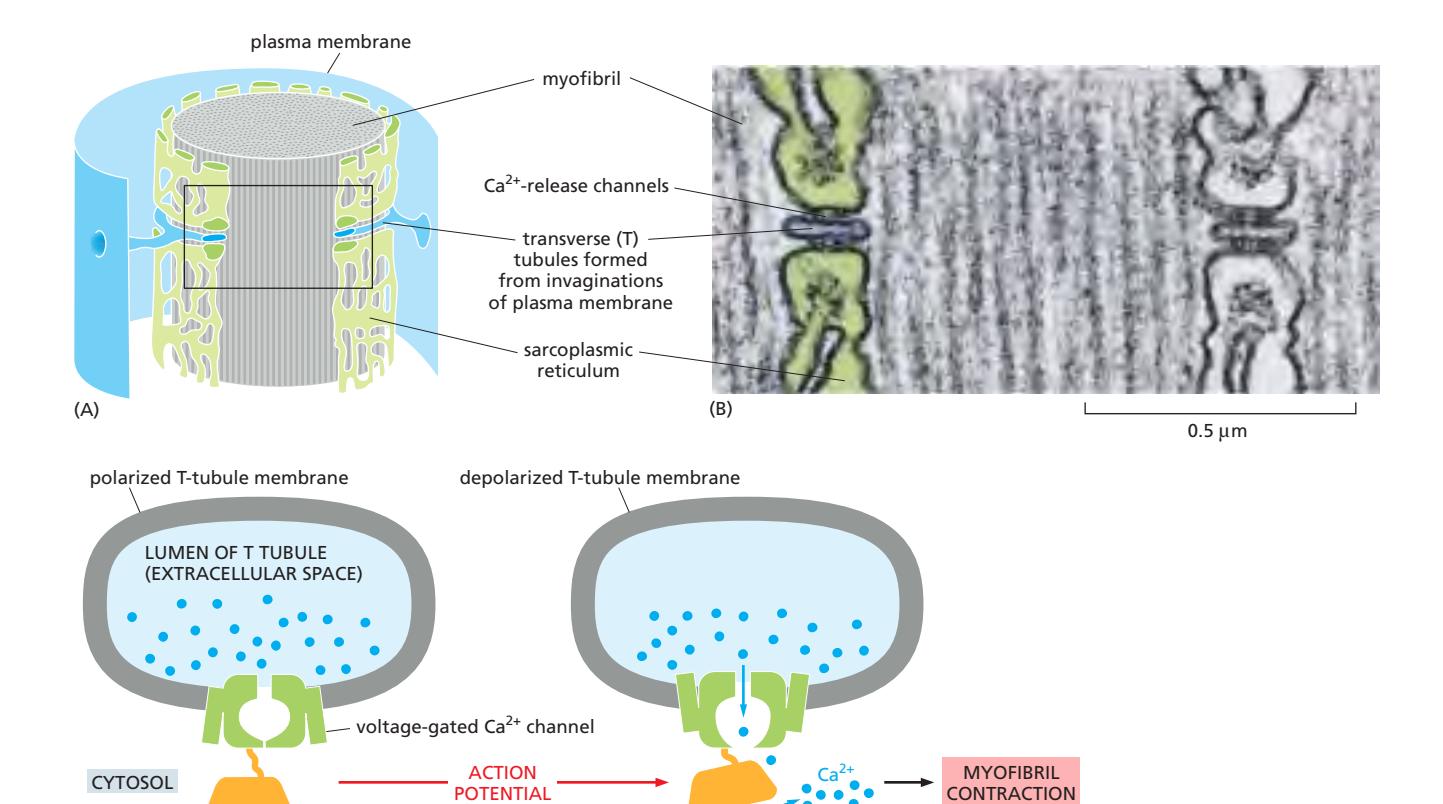
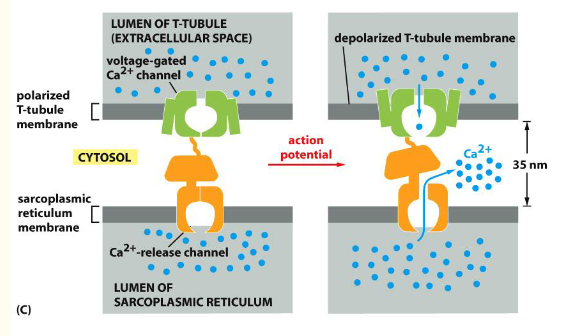 Fig 16–35. T tubule과 sarcoplasmic reticulum. (A) 구조 모식도. (B) T tubule 단면 전자현미경. (C) 전압 개폐 Ca²⁺ channel이 SR의 Ca²⁺-release channel을 여는 과정.
Fig 16–35. T tubule과 sarcoplasmic reticulum. (A) 구조 모식도. (B) T tubule 단면 전자현미경. (C) 전압 개폐 Ca²⁺ channel이 SR의 Ca²⁺-release channel을 여는 과정.
T tubule (transverse tubule): 골격근 세포의 plasma membrane이 안쪽으로 깊게 함입되어 형성된 막 구조로, 각 myofibril을 둘러싼다. 신경에서 오는 action potential이 T tubule을 통해 세포 내부로 빠르게 전파되어 수 ms 내에 모든 sarcomere에 동시 도달한다.
Sarcoplasmic reticulum (SR): 각 myofibril을 망상으로 감싸는 특수화된 ER. T tubule에서 전달되는 전기 신호를 Ca²⁺ 방출로 변환한다.
수축 개시 신호 전달 경로:
- 신경의 action potential → T tubule을 통해 세포 내부로 전파
- T tubule 막의 전압 개폐 Ca²⁺ channel 활성화
- 인접한 SR 막의 Ca²⁺-release channel (ryanodine receptor) 개방
- Ca²⁺가 세포질로 대량 방출
- Ca²⁺가 troponin C에 결합 → myosin binding site 노출 → 수축 시작
이완: SR 막의 **ATP 의존성 Ca²⁺ pump (Ca²⁺-ATPase)**가 Ca²⁺를 SR 내강으로 능동 수송 → 약 30 ms 내에 휴지 수준 복구 → myofibril 이완
ATP 소비 과정:
- 필라멘트 sliding: Myosin II ATPase → actin 필라멘트를 따라 이동하여 수축력 생성
- Ca²⁺ 펌핑: Ca²⁺-ATPase (SR 막) → Ca²⁺를 SR로 재흡수하여 이완 유도
Control of Skeletal Muscle Contraction by Troponin
→ troponin 참조
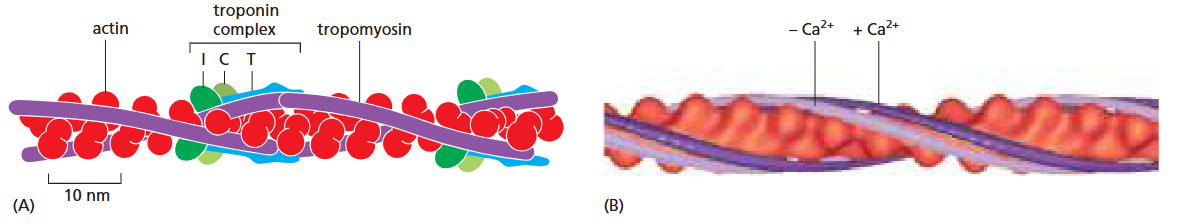 Fig 16–36. Troponin에 의한 골격근 수축 조절. Ca²⁺ 존재(짙은 보라)와 부재(옅은 보라) 시 tropomyosin 위치 변화.
Fig 16–36. Troponin에 의한 골격근 수축 조절. Ca²⁺ 존재(짙은 보라)와 부재(옅은 보라) 시 tropomyosin 위치 변화.
Troponin은 세 가지 폴리펩타이드로 구성된 복합체로, Ca²⁺에 의해 골격근 수축을 조절한다.
| Subunit | 결합 상대 |
|---|---|
| Troponin T (TnT) | Tropomyosin |
| Troponin I (TnI) | Actin + TnT |
| Troponin C (TnC) | Ca²⁺ (최대 4개); calmodulin의 특수화된 형태 |
작동 원리:
- 휴지 상태 (Ca²⁺ 낮음): TnI-TnT 복합체가 tropomyosin을 정상 결합 홈에서 벗어난 위치로 당김 → myosin head의 결합 방해 → 근육 이완 상태 유지
- 수축 개시 (Ca²⁺ 상승): SR에서 방출된 Ca²⁺가 TnC에 결합 → TnI가 actin에 대한 grip 해제 → tropomyosin이 정상 위치로 복귀하여 myosin binding site 노출 → myosin head가 actin 필라멘트를 따라 이동하여 수축 발생