Cell Migration by Actin Cytoskeleton
세포 이동(crawling)은 actin cytoskeleton에 의존하며, 핵심 원리는 leading edge에서의 actin 중합을 통한 membrane 밀어냄과 세포 후방에서의 수축이다. 이동 모드는 actin이 조직화되는 방식에 따라 다르게 나타난다.1
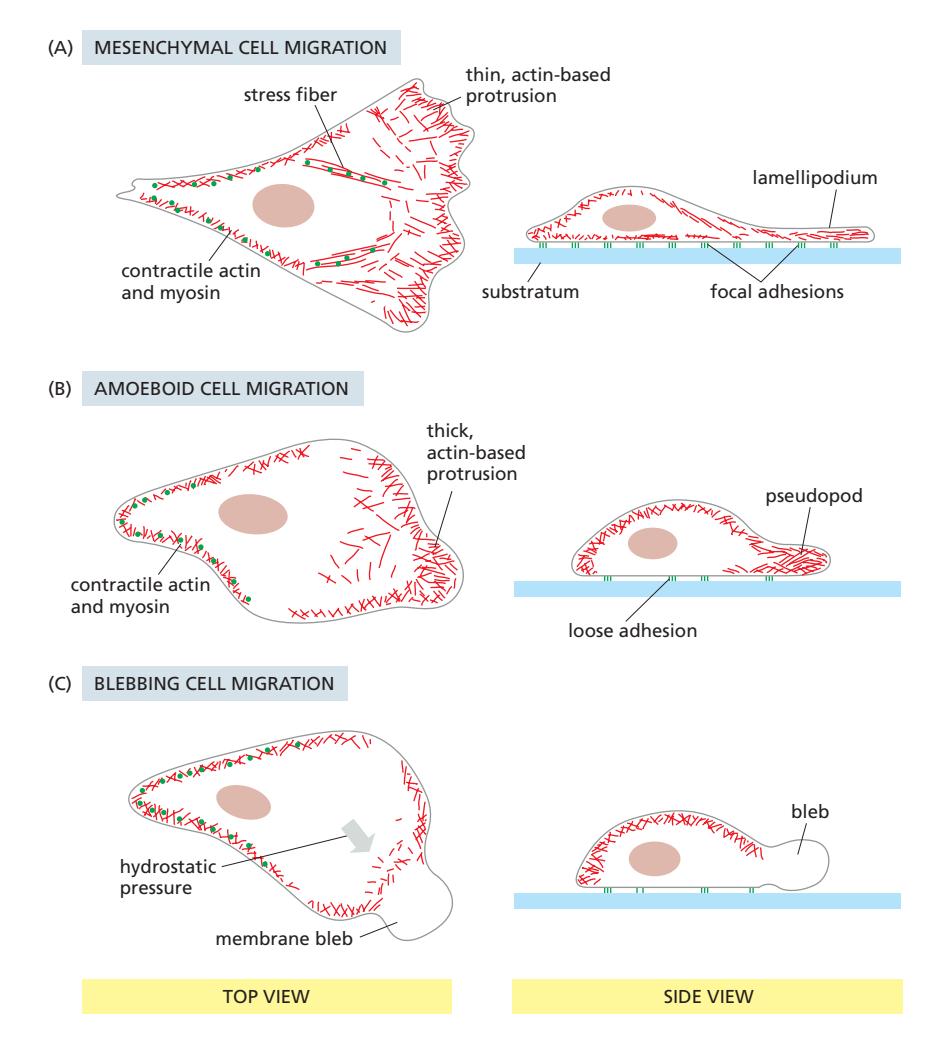 Fig 16–19. 세포 이동의 세 가지 모드. (A) Mesenchymal mode: Arp2/3가 lamellipodia를 만들고 stress fiber가 focal adhesion에 연결. (B) Amoeboid mode: Arp2/3가 3D pseudopod을 만들어 빠른 이동. (C) Blebbing: cortex와 membrane이 분리되어 수압으로 돌출.
Fig 16–19. 세포 이동의 세 가지 모드. (A) Mesenchymal mode: Arp2/3가 lamellipodia를 만들고 stress fiber가 focal adhesion에 연결. (B) Amoeboid mode: Arp2/3가 3D pseudopod을 만들어 빠른 이동. (C) Blebbing: cortex와 membrane이 분리되어 수압으로 돌출.
세 가지 이동 모드
Mesenchymal cell migration은 fibroblast나 유리 표면의 상피세포에서 나타난다. Arp2/3 complex가 leading edge에서 actin 중합을 유도해 lamellipodium을 형성하고, cofilin이 오래된 ADP-actin 필라멘트를 해체한다. 결과적으로 front에서 조립, back에서 분해가 일어나는 패턴이 treadmilling과 유사하다. 이 모드는 integrin 기반 focal adhesion을 통한 기질(ECM) 부착이 필요하며, 속도는 1 μm/min 미만으로 느리다.
Amoeboid cell migration은 중성구 같은 백혈구에서 나타나며 훨씬 빠르다. Arp2/3 complex의 국소 활성화로 두꺼운 3D pseudopod이 폭발적으로 연장된다. Integrin 기반 부착에 덜 의존하며 더 둥근 형태를 유지한다.
Blebbing은 plasma membrane이 actin cortex에서 국소적으로 분리되어 세포 후방의 actin-myosin 수축이 만드는 수압으로 돌출을 형성하는 방식이다. 돌출 후 안쪽 표면에 새 actin cortex가 재형성된다.
lamellipodium의 형성과 이동
Mesenchymal cell migration.
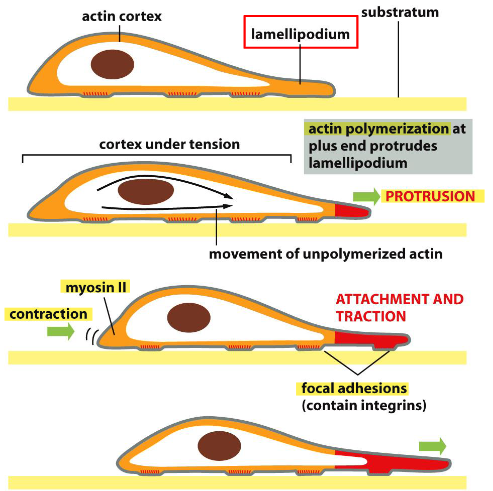
1️. 앞쪽에서 밀기 (Protrusion)
- 액틴 중합 → 막을 앞으로 밀어냄
- lamellipodium 형성
2️. 앞쪽 고정 (Attachment)
- 새 focal adhesion 형성 → 바닥에 붙음
3️. 뒤쪽 끌기 (Contraction)
- 뒤쪽 액틴-마이오신 수축 → 세포 몸통을 앞으로 당김
4️. 뒤쪽 떼기 (Detachment)
- 뒤쪽 adhesion 해체 → 앞으로 이동 가능
Protrusion of the Actin Meshwork at the Leading Edge
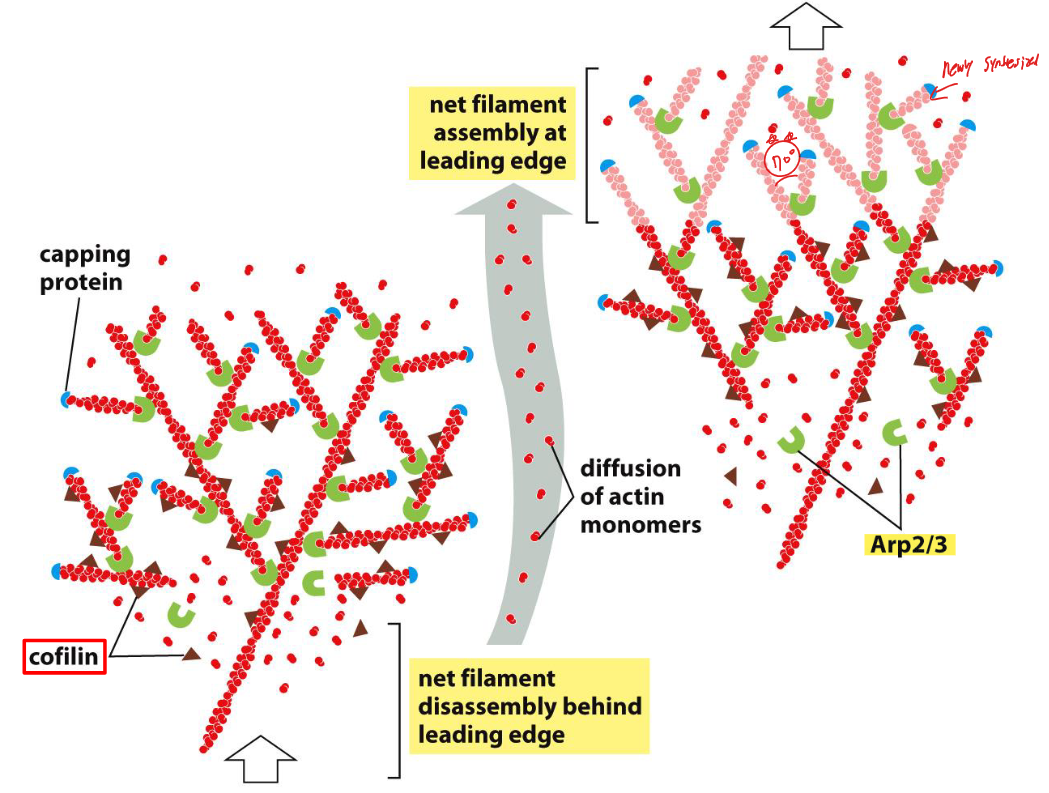 Fig 16–80. Leading edge에서 actin meshwork protrusion 모델. 두 시점에서 lamellipodium 전진을 보여주며, 나중 시점의 새로 조립된 구조는 더 밝은 색으로 표시.
Fig 16–80. Leading edge에서 actin meshwork protrusion 모델. 두 시점에서 lamellipodium 전진을 보여주며, 나중 시점의 새로 조립된 구조는 더 밝은 색으로 표시.
Leading edge에서 actin meshwork가 앞으로 나아가는 메커니즘은 다음 단계로 이루어진다.
- Nucleation: Arp2/3 complex가 leading edge front에서 기존 필라멘트 측면에 새 필라멘트를 nucleate한다. 주로 70° 각도로 가지를 뻗는다.
- Elongation: 새로 생긴 필라멘트의 plus end가 길어지면서 plasma membrane을 앞으로 민다. 이 힘이 가능한 이유는 network 뒤쪽에 어떤 형태의 고정(anchorage)이 있기 때문이다.
- Capping: 일정한 속도로 plus end가 capping protein에 의해 막힌다.
- Depolymerization: 새로 중합된 subunit이 filament 내에서 ATP를 ADP로 가수분해하면, cofilin이 ADP-actin 필라멘트를 선택적으로 인식하여 depolymerization을 가속한다 (→ treadmilling 참조).
이 과정의 결과로 front에서는 net 조립, rear에서는 net 분해가 공간적으로 분리된다. 따라서 actin 필라멘트 network 전체는 앞으로 전진하지만, 개별 필라멘트는 기질에 대해 정지 상태를 유지한다.
Cofilin이 분해시키지 못한 일부 actin은 lamellipodium 뒤쪽에 남아, myosin II와 함께 이후 이동 단계에 기여한다.
관련 단백질의 기능에 대한 자세한 내용은 major accessory proteins of the actin cytoskeleton 참조.
Footnotes
-
2023 기말 23번 — 6종류 actin array(lamellipodia, filopodia, stress fiber, cortical actin network 등)와 세포 이동(mesenchymal/amoeboid 모드)에서의 역할이 정답 근거로 활용됨. ↩