Centrosome
Centrosome은 동물 세포에서 가장 중요한 MTOC (microtubule-organizing center)로, 핵 옆 세포질에 위치한다.1 Minus end를 고정하고 plus end를 세포 바깥쪽으로 향하게 하여 microtubule을 nucleate하며, 세포 내 organelle 배치의 전체적인 좌표계를 제공한다.2
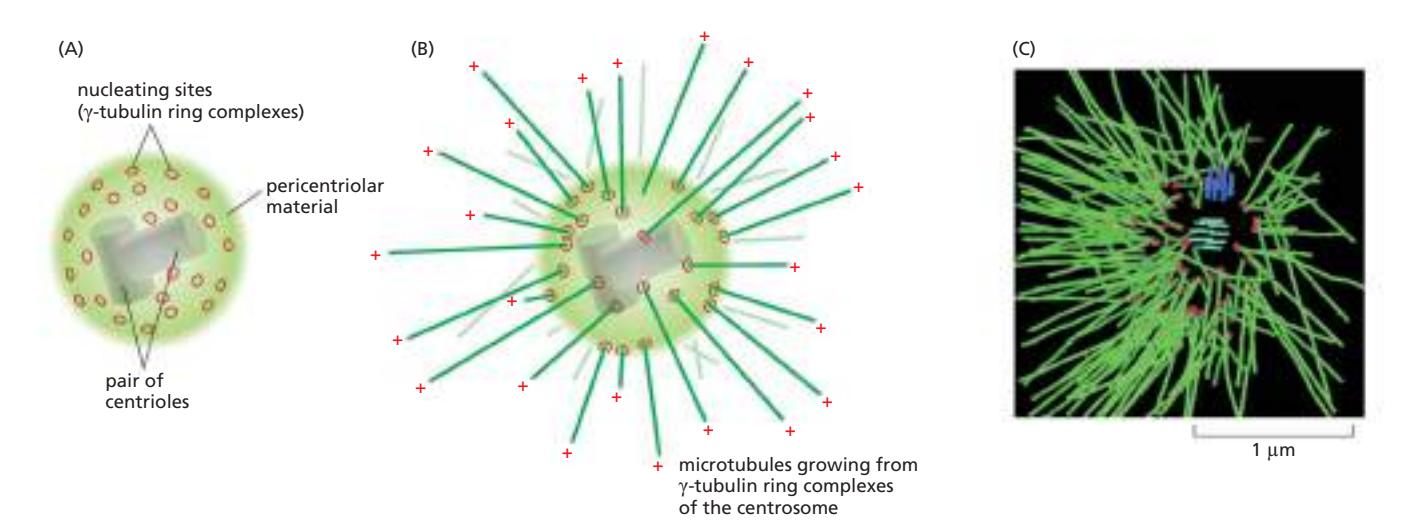 Fig 16–42. Centrosome의 구조. (A) 핵 옆에 위치하며, centriole 쌍을 pericentriolar material이 둘러싸는 구조. (B) Microtubule이 부착된 centrosome. Minus end는 centrosome에 묻히고 plus end는 세포질 쪽으로 자유롭게 뻗는다. (C) C. elegans 세포 MTOC에서 방사상으로 뻗은 microtubule thicket.
Fig 16–42. Centrosome의 구조. (A) 핵 옆에 위치하며, centriole 쌍을 pericentriolar material이 둘러싸는 구조. (B) Microtubule이 부착된 centrosome. Minus end는 centrosome에 묻히고 plus end는 세포질 쪽으로 자유롭게 뻗는다. (C) C. elegans 세포 MTOC에서 방사상으로 뻗은 microtubule thicket.
γ-Tubulin Ring Complex (γ-TuRC)
Microtubule nucleation의 핵심은 **γ-tubulin ring complex (γ-TuRC)**이다.
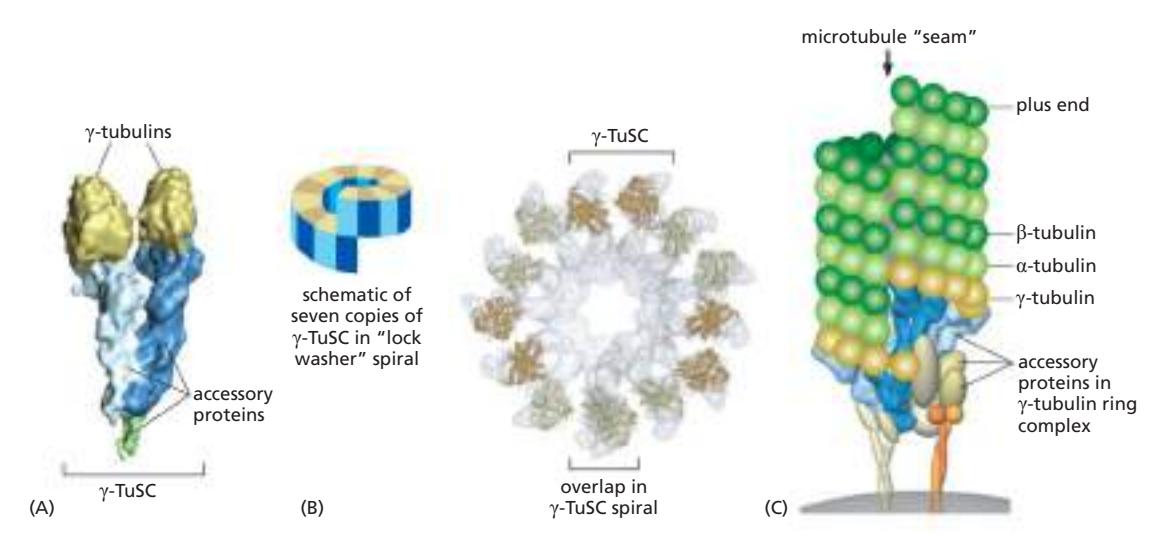 Fig 16–41. γ-TuRC. (A) γ-tubulin 2개와 accessory protein 쌍으로 구성된 γ-TuSC. (B) 7개의 γ-TuSC가 나선형으로 모여 13개의 γ-tubulin이 원형으로 노출. (C) γ-TuRC가 microtubule minus end를 nucleate하는 모델.
Fig 16–41. γ-TuRC. (A) γ-tubulin 2개와 accessory protein 쌍으로 구성된 γ-TuSC. (B) 7개의 γ-TuSC가 나선형으로 모여 13개의 γ-tubulin이 원형으로 노출. (C) γ-TuRC가 microtubule minus end를 nucleate하는 모델.
γ-tubulin 두 개가 한 쌍의 accessory protein과 결합하여 γ-TuSC (γ-tubulin small complex)를 형성한다. 7개의 γ-TuSC가 모이면 나선형 구조를 이루어 13개의 γ-tubulin이 원형으로 노출되고, 이것이 microtubule의 13 protofilament와 일치하는 주형을 제공하여 minus end를 nucleate한다. 나선형 배열로 인해 두 protofilament 사이에 하나의 longitudinal seam이 생기는 것이 특징이다. microtubule합성은 plus end에서 일어나고, minus end에서는 고정만 한다.
Pericentriolar Material (PCM)
Centriole 주변을 감싸는 PCM (pericentriolar material)은 섬유성 단백질들의 무정형 망상 구조로, γ-TuRC가 여기에 묻혀 microtubule nucleation이 일어난다. PCM은 biomolecular condensation 과정을 통해 형성되는 dense spherical matrix로 생각된다.
Centriole
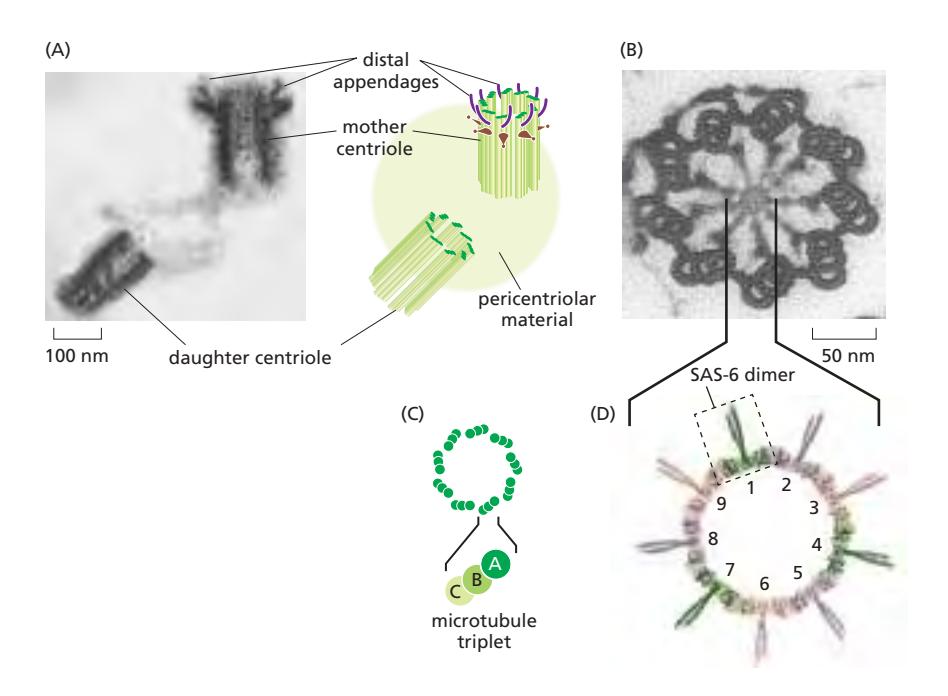 Fig 16–43. Centriole 쌍. (A) Distal appendage를 가진 mother centriole과 인접한 daughter centriole 단면. (B) Protozoan centriole 단면: 9세트의 microtubule triplet이 원통형 배열. (C) Triplet 구성. (D) SAS-6 단백질이 cartwheel의 9중 대칭을 형성.
Fig 16–43. Centriole 쌍. (A) Distal appendage를 가진 mother centriole과 인접한 daughter centriole 단면. (B) Protozoan centriole 단면: 9세트의 microtubule triplet이 원통형 배열. (C) Triplet 구성. (D) SAS-6 단백질이 cartwheel의 9중 대칭을 형성.
- Centrosome 내부에는 L자형으로 배치된 한 쌍의 centriole이 있다.
- 각 centriole은 9세트의 microtubule triplet이 원통형으로 배열된 구조로 9중 대칭을 갖는다.
- 각 triplet은 하나의 완전한 microtubule (A microtubule)에 두 개의 불완전한 microtubule (B, C microtubule)이 융합된 구조다.
- 중앙 cartwheel 구조의 9중 대칭은 SAS-6 단백질이 coiled-coil dimer를 이루어 9개가 자기조립하여 링을 형성함으로써 만들어진다.
Mother vs. Daughter Centriole
한 쌍의 centriole은 기능적으로 구분된다.
| Mother centriole | Daughter centriole | |
|---|---|---|
| 형성 시기 | 이전 세포주기 | 현재 S phase |
| 구조적 특징 | Distal appendage를 가짐 | Appendage 없음 |
| 기능적 차이 | Primary cilia의 basal body로 기능 | 분열 후 새로운 mother가 됨 |
Centrosome 복제와 세포 분열
Centrosome은 S phase에 복제되어 각 centriole 옆에 새로운 딸 centriole이 형성된다. Mitosis가 시작되면 두 centrosome이 분리되어 mitotic spindle의 양극을 형성한다. 분열이 끝나면 각 딸세포는 하나의 centrosome을 갖게 되며, 그 안에 한 쌍의 centriole이 들어 있다.
Centrosome의 이중 기능: Primary Cilia의 Basal Body
Interphase에서 mother centriole은 세포막 근처로 이동하여 primary cilia의 basal body로 기능한다.
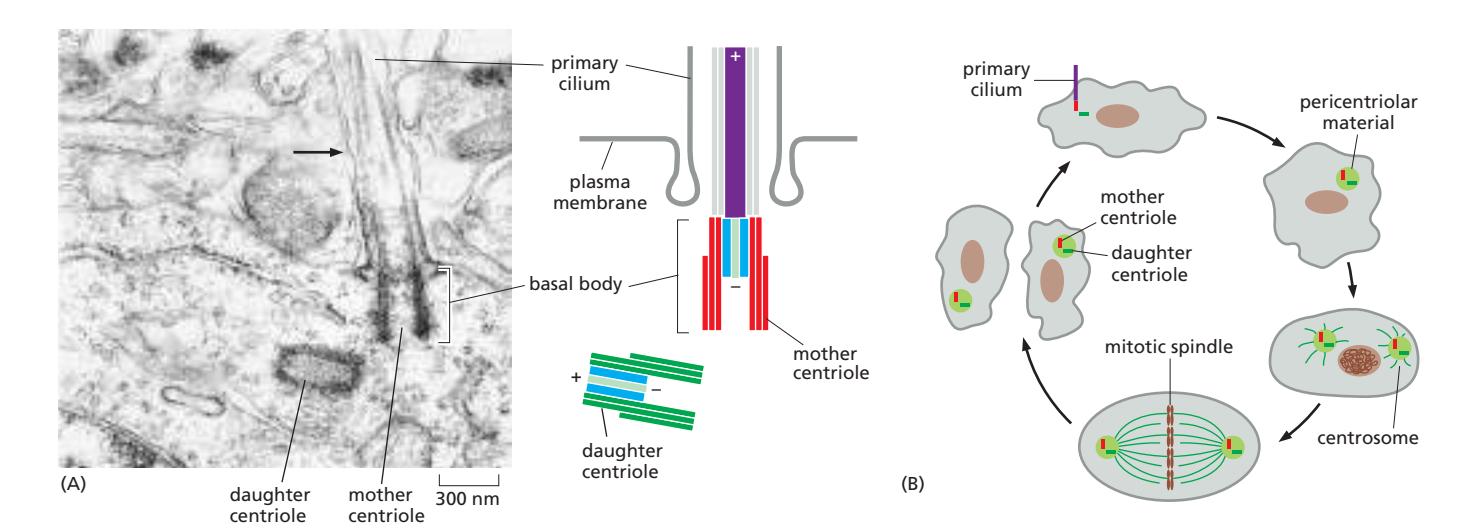 Fig 16–61. Primary cilia. (A) Mouse neuron primary cilium의 basal body. Mother centriole이 basal body가 되어 axoneme nucleation을 담당. (B) Centriole의 기능적 전환 — interphase에서는 primary cilium의 basal body, 분열기에는 mitotic spindle의 centrosome.
Fig 16–61. Primary cilia. (A) Mouse neuron primary cilium의 basal body. Mother centriole이 basal body가 되어 axoneme nucleation을 담당. (B) Centriole의 기능적 전환 — interphase에서는 primary cilium의 basal body, 분열기에는 mitotic spindle의 centrosome.
이 전환 과정은 세포 주기와 연동된다.
- 분열 후 (Interphase): Mother centriole이 세포막 근처로 이동 → primary cilium 형성
- 분열 준비 (S phase): Primary cilium 흡수 → centriole 복제
- 분열기 (Mitosis): 두 centrosome이 spindle 양극 형성
관련 노트
- structure of a microtubule — α/β-tubulin heterodimer, 13 protofilament 구조
- preferential growth of microtubules — plus end 위주 성장, dynamic instability
- axoneme — centriole에서 형성되는 axoneme 구조
- Microtubule organization in a neuron — 신경 세포에서의 microtubule 극성
Footnotes
-
2025 기말 18번 — Centrosome이 핵막 안이 아닌 세포질(핵 바깥)에 위치한다는 내용이 정답 ②의 근거로 활용됨. ↩
-
2023 기말 22번 — γ-TuRC가 microtubule minus end를 nucleate하는 주형을 제공한다는 내용이 정답 근거로 활용됨. ↩