Chapter 13-2: Intracellular Membrane Traffic 정리본
6주차 강의노트 기반 | 출처: chapter 13 강의 II 2026.pdf
1. Lysosome
Lysosome은 세포 내 소화의 주요 장소이다.
구조적 특징
- 내부 pH ~4.5–5.0 유지 (강산성)
- **V-type ATPase (H⁺ pump)**가 ATP hydrolysis 에너지로 H⁺를 lumen으로 펌핑
- 막 단백질은 대부분 highly glycosylated → lysosomal protease로부터 보호
- Transport 단백질: 분해 산물(amino acid, sugar, nucleotide)을 세포질로 운반
Lysosomal Hydrolase
- 산성 환경(pH 4.5–5.0)에서 최적 활성
- Hydrolase가 cytosol로 방출되더라도 중성 pH(~7.2)에서는 활성이 낮아 세포 손상 제한
- 종류: protease, lipase, nuclease, glycosidase 등
Lysosome의 형태적 다양성
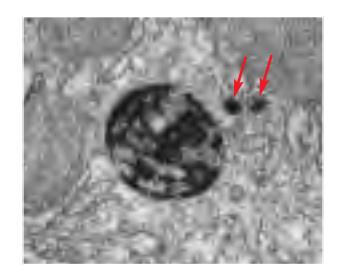
 전형적인 “classical” lysosome은 내용물이 거의 소화된 후 남은 잔류물만 포함한 상태이다.
late endosome은 lysosomal hydrolase를 받거나lysosome과 융합하여 내용물을 가수분해한다.
가수분해가 끝나고 enzyme만 남으면 비로소 lysosome이 된다.
이처럼 lysosome과 late endosome은 구별하기 쉽지 않고, 거의 같은 기관이라 보아도 무방하다.
전형적인 “classical” lysosome은 내용물이 거의 소화된 후 남은 잔류물만 포함한 상태이다.
late endosome은 lysosomal hydrolase를 받거나lysosome과 융합하여 내용물을 가수분해한다.
가수분해가 끝나고 enzyme만 남으면 비로소 lysosome이 된다.
이처럼 lysosome과 late endosome은 구별하기 쉽지 않고, 거의 같은 기관이라 보아도 무방하다.
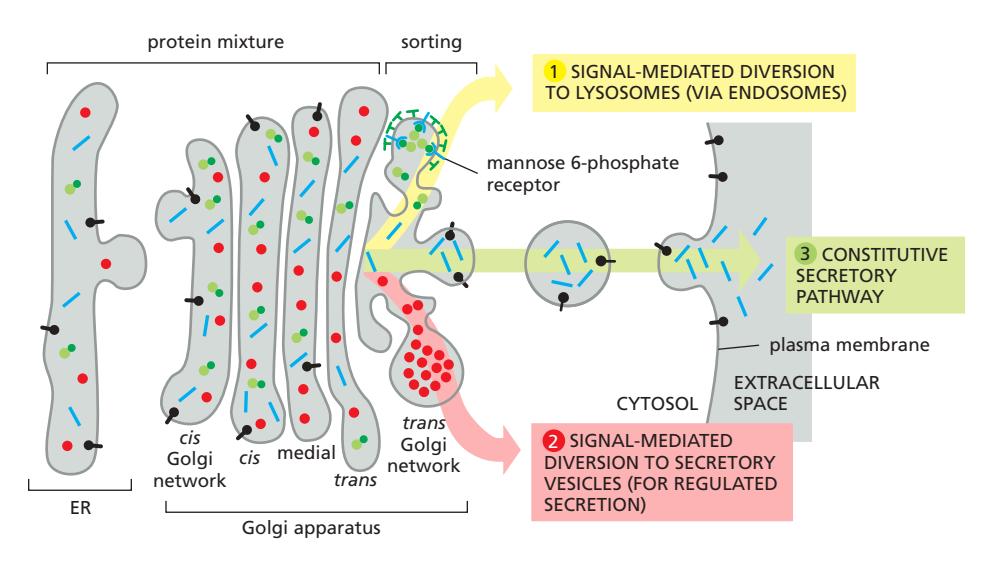
2. Lysosomal Hydrolase 수송: M6P Receptor System
M6P receptor system은 TGN에서 lysosomal hydrolase를 endosome으로 전달하는 가장 잘 이해된 sorting 메커니즘이다.
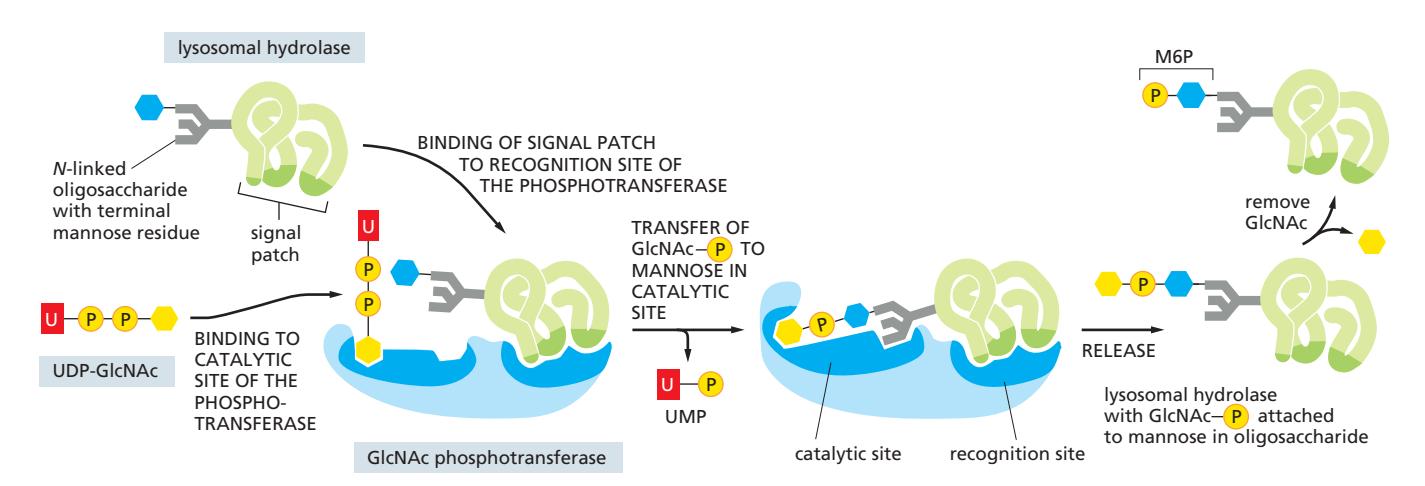
M6P Tag의 부착
- Cis Golgi network에서 GlcNAc phosphotransferase가 lysosomal hydrolase의 N-linked oligosaccharide를 인식
- Two-step modification → Mannose-6-phosphate (M6P) 노출
- 각 hydrolase에 multiple M6P tag 부착 → M6P receptor에 high affinity
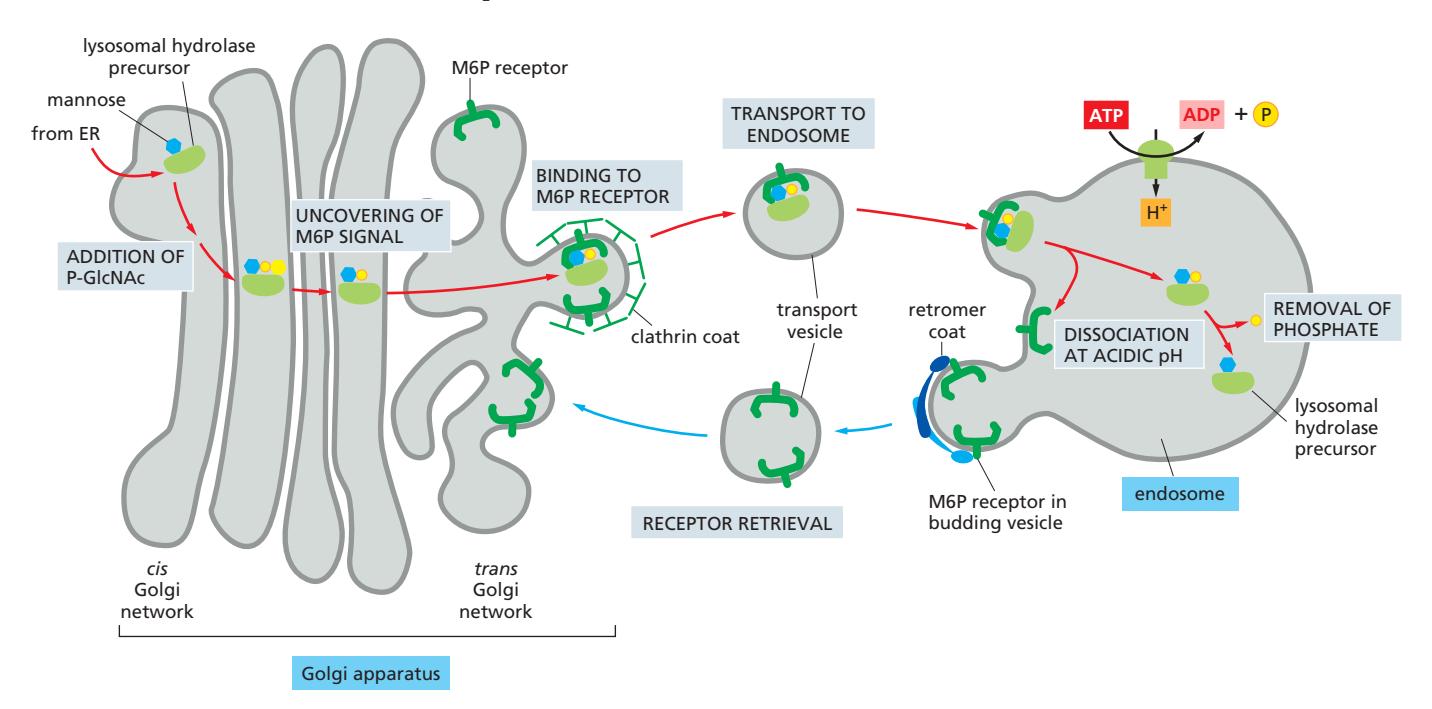
M6P Receptor의 역할
- Transmembrane protein (TGN에 위치)
- Lumen 쪽에서 M6P group 인식 → lysosomal hydrolase 결합
- Cytosolic 쪽에서 clathrin coat의 adaptor protein 결합
- Clathrin-coated vesicle로 hydrolase packaging
pH-dependent 결합
| 구획 | pH | M6P receptor 상태 |
|---|---|---|
| TGN | 6.5–6.7 | M6P에 강하게 결합 |
| Early endosome | ~6.0 | M6P로부터 방출 |
Receptor 재활용
- Endosome의 낮은 pH → hydrolase가 receptor에서 분리
- M6P에서 phosphate 제거 (hydrolase가 receptor와 함께 TGN으로 돌아가지 않도록)
- 빈 M6P receptor → retromer-coated vesicle로 회수 → TGN 복귀
- M6P receptor 재사용
KDEL receptor (COPI vesicle) vs. M6P receptor (clathrin/retromer vesicle): 같은 pH-sensitive 원리이지만 다른 coat 사용
Lysosomal Storage Disease
I-cell Disease (Inclusion-cell Disease)
I-cell disease는 가장 심한 형태의 lysosomal storage disease이다:
- 원인: GlcNAc phosphotransferase 결함 → M6P tagging 불가
- 결과: Lysosomal hydrolase가 lysosome으로 배달되지 않고 혈액으로 분비됨
- Lysosome에 거의 모든 hydrolytic 효소 결여
- 소화되지 않은 기질이 축적 → lysosome이 큰 inclusion 형성 (효소 없는 쓰레기 vesicle)
- 모든 기관계 영향, 개인은 6–7세를 넘기지 못함
- Hepatocyte는 M6P-independent pathway를 가져 예외적으로 정상 작동
Hurler’s Disease
Hurler’s disease는 I-cell disease와 달리 개별 lysosomal hydrolase의 structural gene mutation으로 발생:
- 특정 유형의 glycosaminoglycan (GAG) chain을 분해하는 데 필요한 lysosomal 효소가 결함적이거나 결여됨
- 해당 효소만 기능하지 않아 GAG 분해물이 lysosome에 축적
- I-cell disease와 달리 특정 효소 하나의 결함 (M6P tagging 자체는 정상)
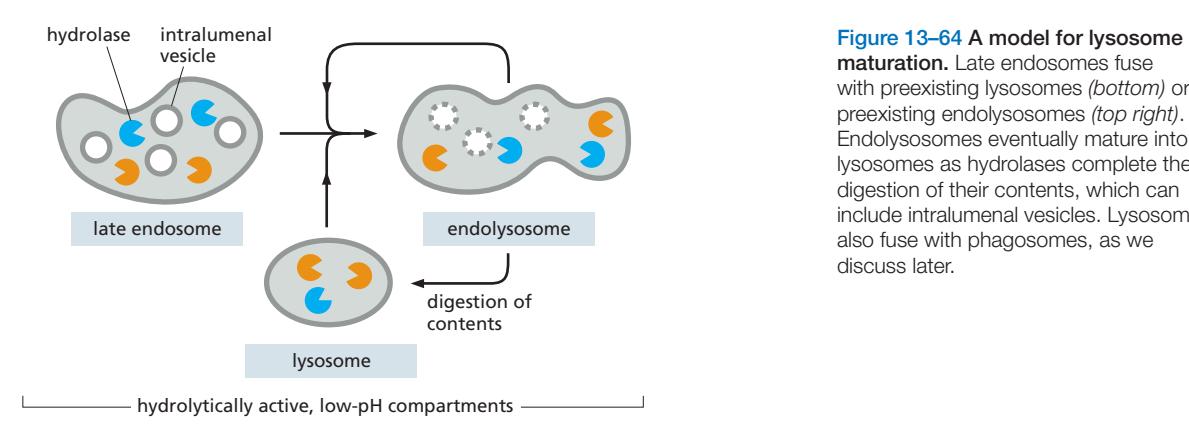
3. Endosome Maturation
Endosome maturation은 early endosome이 시간이 지나면서 late endosome으로 변화하는 과정이다.
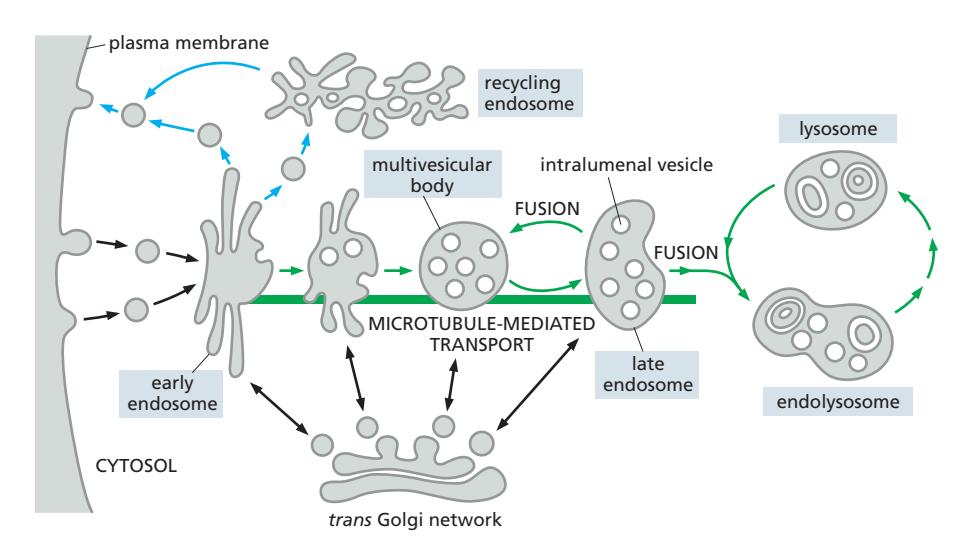
과정
Early endosome (pH ~6.0):
- Tubular + vacuolar domain 보유
- 약 10분간 들어오는 endocytic vesicle 수용
- Tubular domain: 대부분의 membrane 표면 / Vacuolar domain: 대부분의 부피
Maturation 단계별 변화:
- 형태 변화: Tubular domain이 plasma membrane으로 재활용 / Vacuolar domain 수정
- 이동: Dynein motor에 의해 microtubule을 따라 핵 쪽으로 이동
- Lysosome 단백질 전달: TGN에서 새로 합성된 lumenal hydrolase + V-type ATPase 전달
- Acidification: V-type ATPase가 H⁺ pump → organelle이 더욱 산성화 → hydrolase 점점 활성화
- Intralumenal vesicle 형성: Endocytosed signaling receptor가 내부 vesicle로 격리 → receptor signaling 정지
Rab5→Rab7 전환Rab Cascades
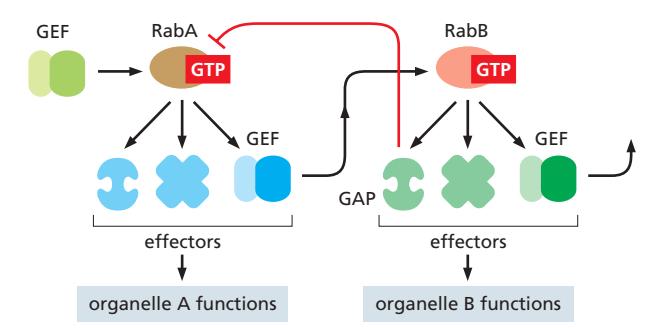
- 시간이 지남에 따라 Rab5-associated patch → Rab7-associated patch로 교체
- Early endosome (Rab5) → Late endosome (Rab7)
- Rab7 effector 세트가 Rab5 effector와 달라 organelle 성질 변화
- 일방향적이고 비가역적: Rab의 자기증폭 특성으로 역전 불가
PI(3)P에서 PI(3,5)P₂으로 전환
하면서 late endosome으로 성숙
Lysosome-Late Endosome Cycle
Lysosome은 late endosome과 융합하여 endolysosome이 되고, 분해가 끝나면 다시 lysosome이 되는 cycle 형성.
따라서 lysosome과 late endosome/endolysosome은 명확히 구분하기 어려운 연속적인 변화 과정.
- 내용물이 분해되는지 여부는 pH 차이에 따라 결정
- lysosomal hydrolase는 late endosome과 lysosome으로 모두 전달됨.
4. Multivesicular Body (MVB)와 ESCRT
ESCRT complex가 intralumenal vesicle (ILV) 형성을 담당한다.
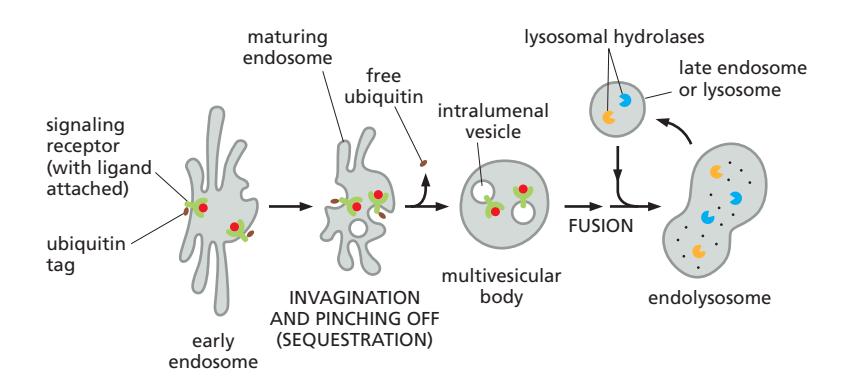
Multivesicular Body
Endosome이 성숙함에 따라:
- 막의 patch가 endosome lumen으로 invaginate
- Pinch off하여 intralumenal vesicle (ILV) 형성. 특정 상황에서 이 ILV를 내보내는 경우 Exosome이라고도 불림
- 세포간 신호로 작용하거나 약물 전달 경로에 이용되기도 함
- Receptor의 cytosolic face의 signal을 차단하기 위함
- 이러한 endosome을 **Multivesicular Body (MVB)**라고 함
- ESCRT (Endosomal Sorting Complex Required for Transport) protein complex에 의해 생성
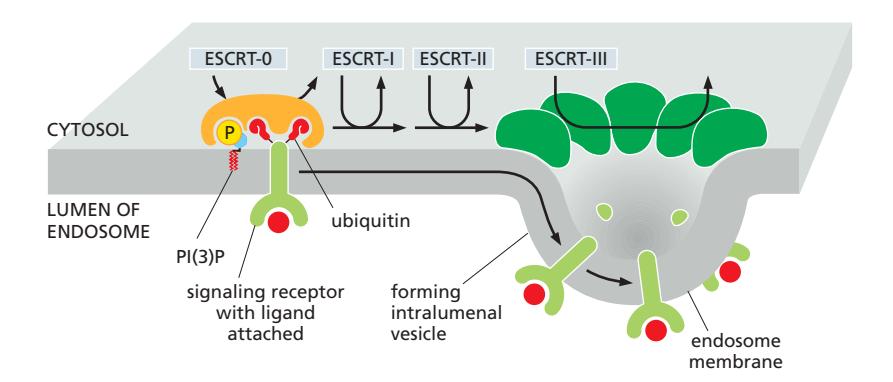
ESCRT의 순차적 작용
- ESCRT-0: Ubiquitin tag + PI(3)P 동시 인식 → 막에 처음 결합, cargo를 다음으로 전달
- ESCRT-I: cargo 받아 ESCRT-II로 전달
- ESCRT-II: cargo를 특정 막 영역에 농축, ESCRT-III 모집
- ESCRT-III: 막에 multimeric assembly 형성 → 막 bend → invagination 매개
ESCRT docking 요구사항: PI(3)P + ubiquitylated cargo 단백질 둘 다 필요
Ubiquitin 제거: Intralumenal vesicle이 닫히기 전에 ubiquitin marker를 제거 → 세포질로 반환하여 재사용
막 굴곡의 방향
ESCRT가 추진하는 ILV budding은 막의 cytosolic 표면에서 멀어지는 방향으로 발생
- 이는 Clathrin coat의 방향과 반대 (clathrin은 cytosolic 쪽으로 구부림)
Late endosome → lysosome 융합 후 ILV 내 ubiquitinated receptor 분해
5. Retrieval Pathway (Endosome에서)
Early endosome의 tubular extension에서 transport vesicle이 budding:
- 선택된 막 단백질 회수 → plasma membrane으로 반환
- 직접 반환 또는 recycling endosome을 거쳐 반환
6. Endocytosis의 종류
Endocytosis는 plasma membrane 성분, fluid, 용질, 거대분자를 흡수하는 과정이다.
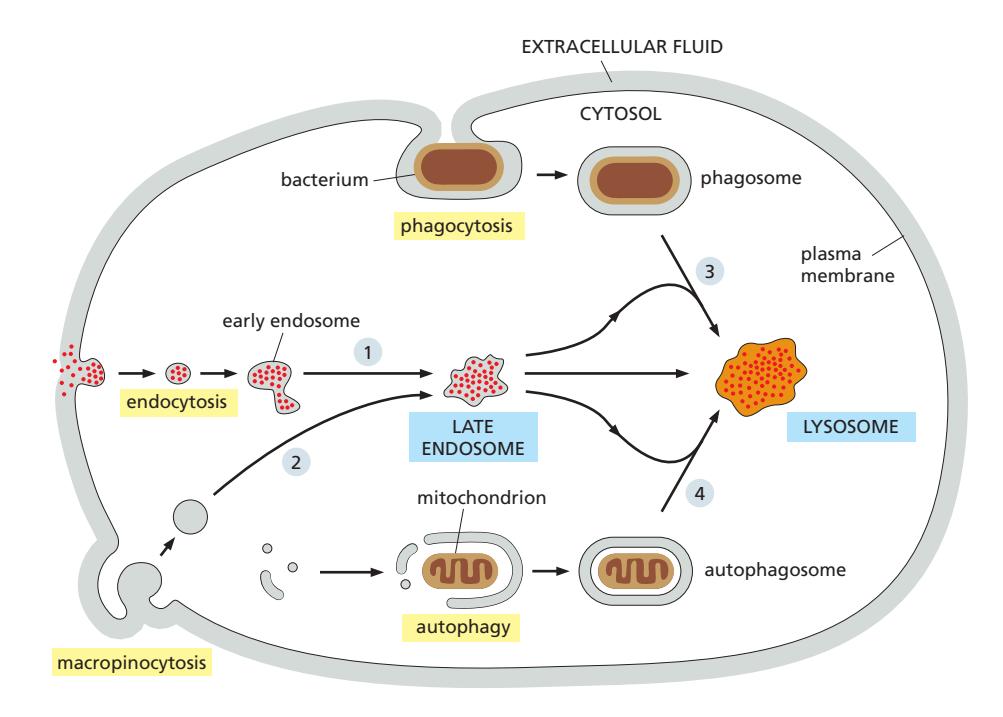
Pinocytosis의 종류
(1) Clathrin-mediated Endocytosis
- 가장 흔한 형태
- Clathrin-coated pit → coated vesicle → coat shed → early endosome 융합
(2) Caveolar Endocytosis
Caveolae는 clathrin-independent membrane invagination이다.
- Caveolae: flask 모양의 invagination
- 구성: Cholesterol + Glycosphingolipid + GPI-anchored protein → lipid raft 유형
- 이들은 lipid raft 형태로 모여있고, 이 모임이 curvature를 만들고, vesicle formation에 필요한 지질이나 protein을 모아놓기 때문에 vesicle 형성에 유리하다. lipid raft라해서 유동성이 낮아지는 것이 아니다.
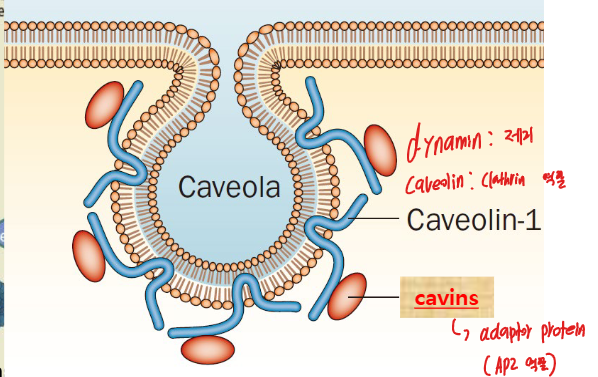
- Caveolin (coat protein): 특이한 integral membrane protein; hydrophobic loop을 cytosolic 쪽에서 막에 삽입 (막을 가로지르지 않음) clathrin과 차이점이 바로 막단백질이라는 점.
- Cavin (adaptor protein.비유하자면.): caveolin에 결합하여 구조 안정화
- 일반적으로 정적 구조 (extra plasma membrane의 저장소)
- 기계적 힘 가해질 시: cavin scaffold 빠른 disassembly → 막 표면적 일시적 증가
- 이는 세포내 actin filament의 stress fiber와 caveolae가 연결되어 있기 때문
(3) Macropinocytosis
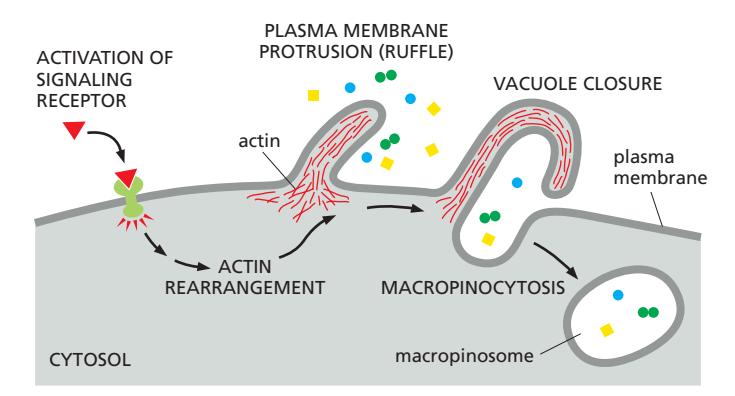
- Macropinocytosis: plasma membrane이 돌출하여 주변 세포외 fluid를 macropinosome으로 둘러싸는 과정
- Clathrin-independent endocytosis
- 비선택적 과정: 세포외 fluid와 그 안의 거대분자·입자를 비특이적으로 trap
- Macropinosome에 receptor가 포함되지 않음 → receptor recycling 불필요
- Actin reorganization으로 membrane ruffling이 일어나며 fluid를 삼킴
- exosome이 macropinocytosis에 의해 macropinosome안에 포함되고 cytosol에서 exosome의 내용물이 방출됨.
형성 과정:
- Cell-surface receptor 활성화 (growth factor, integrin ligand 등). 이 receptor는 macropinosome에 포함되지 않음.
- Actin polymerization → membrane ruffle 형성
- Ruffle의 끝이 서로 또는 세포막과 융합 → macropinosome 형성
분해 경로:
- Macropinosome은 재활용 없이 오직 분해만 진행
- Acidification → late endosome 또는 endolysosome과 융합
- Ras oncogene 활성화에 의해 자극 → 암세포에서 증가 (영양소 획득)
Phagocytosis
→ phagocytosis 참조
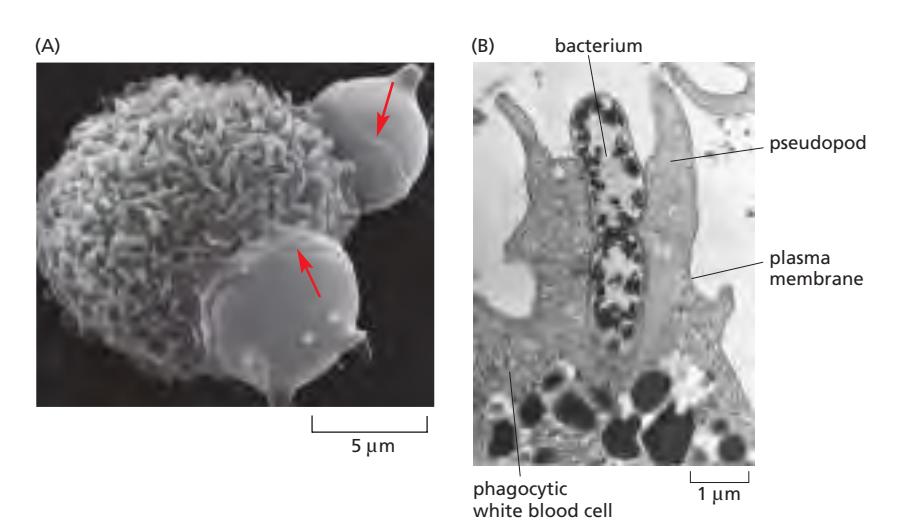
Phagocytosis(“cell eating”)는 면역 세포가 phagosome이라는 큰 endocytic vesicle을 이용하여 큰 입자(미생물, 죽은 세포 등)를 섭취하는 endocytosis의 특수한 형태이다. 면역세포 특이적인 현상이다.
수행 세포: Macrophage와 Neutrophil
Opsonization: 항원에 항체가 결합 → Fc receptor가 항체의 Fc 부위 인식 → phagocytosis 유도 → 항원들을 한꺼번에 제거
Phagosome의 운명:
- Phagosome → lysosome 융합 → 내용물 분해
- 소화되지 않는 물질 → residual body 형성 → exocytosis로 세포 외 배출
- Phagosome 크기는 섭취된 입자 크기에 따라 결정됨 (세포 자체만큼 거의 클 수 있음)
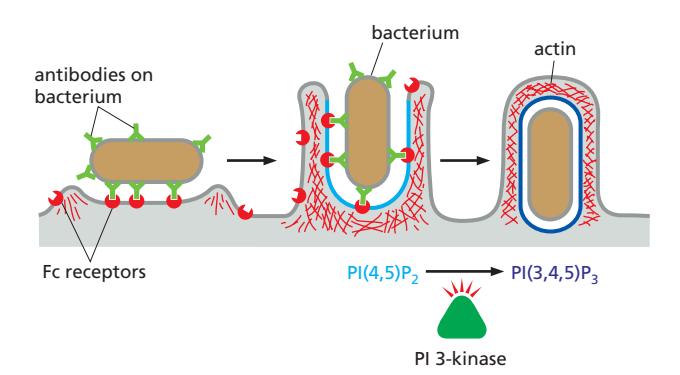
메커니즘 (cargo-triggered process):
 하이라이트만 외우기
하이라이트만 외우기
- Cell-surface receptor(Fc receptor) 활성화 필요; 입자가 먼저 phagocyte 표면에 결합해야 함
- FcR clustering → Syk → PIP5K 활성화 → PI(4,5)P₂ 생성 → WASP → Arp2/3 → actin reorganization → pseudopod 형성 동시에 Syk → PI3K → PI(3,4,5)P₃ 생성 → Rac1 추가 활성화 (양성 피드백)
- PI(4,5)P₂ (cup 전체): actin branching → pseudopod 확장 PI(3,4,5)P₃ (cup 가장자리): 추가 actin 중합 → cup이 타겟 감쌈
- Tip에서 SHIP1 활성화 → PI(3,4,5)P₃ 소멸 → 신호 꺼짐 → actin depolymerization → closure → PI(3)P 생성 → phagosome으로 maturation “Don’t-eat-me” / “Eat-me” Signal:
- 살아있는 세포: cell-surface protein이 macrophage의 inhibitory receptor에 결합 → phagocytosis 억제
- Apoptotic 세포: “Eat-me” signal 획득 (예: 외부로 노출된 phosphatidylserine) + “Don’t-eat-me” signal 상실 → macrophage에 의해 신속히 phagocytose됨
7. Autophagy
→ Autophagy 참조
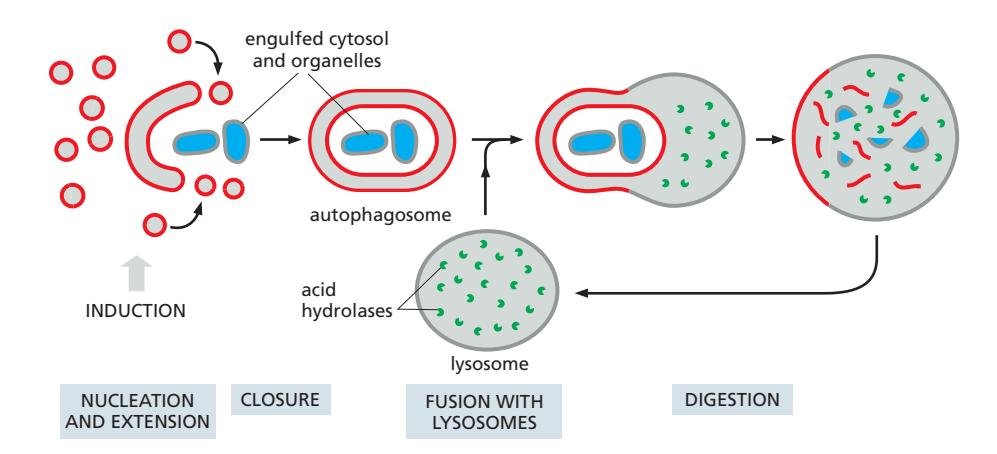
Autophagy(“self-eating”)는 세포질의 일부가 autophagosome이라는 이중막 구조로 포획되어 lysosome과 융합한 뒤 내용물이 분해되는 과정이다.
종류
| 종류 | 조건·대상 | 특성 |
|---|---|---|
| Nonselective autophagy | Starvation 상태 | 아미노산·에너지원 확보를 위해 cytoplasm bulk portion을 무작위 격리·분해 |
| Selective autophagy | 표식(ubiquitin) 붙은 특정 cargo | Mitophagy(손상 미토콘드리아), xenophagy(침입 미생물) 등 |
Autophagosome 조립 과정
- 시작: 이상 신호 전달(mTOR 억제; starvation 상황. nonselective autophagy)
- 막 형성 (Nucleation & Extension): vesicle 융합 → 납작한 disc 형성 → cup-shaped 구조로 말림 → cup 끝이 서로 융합 → 이중막이 내용물 포획
- Lysosome 융합 (Fusion): autophagosome 외막이 SNARE-mediated process로 lysosome과 융합
- 분해 (Digestion): 내막과 포획된 cargo가 lysosomal hydrolases에 의해 가수분해됨
이중막 구조의 의미:
- 외막: lysosome과 융합 (intact 상태)
- 내막: lysosome 내로 방출된 후 분해됨
Mitophagy
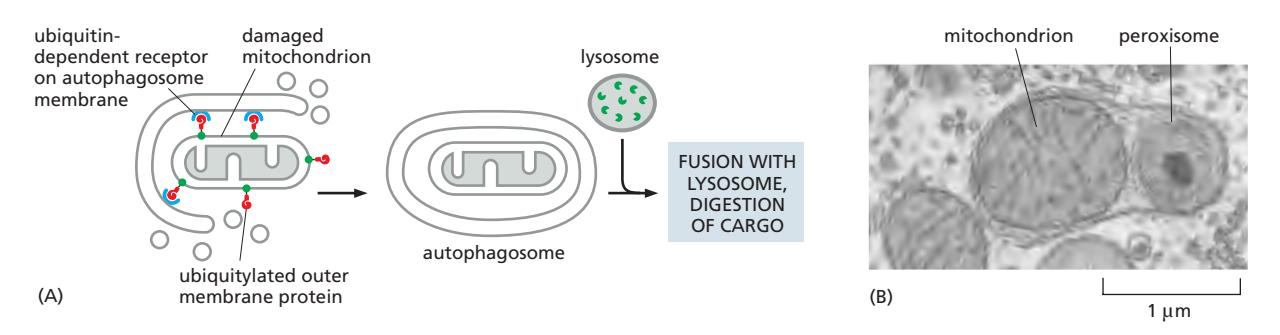
손상된 mitochondria의 selective autophagy:
Selective Autophagy 메커니즘
Cargo-specific receptor가 분해될 cargo를 forming autophagosome의 오목한 막 표면으로 모집. 대부분 ubiquitin이 cargo 표식에 사용됨.
8. Receptor-mediated Endocytosis: LDL 사례
Receptor-mediated endocytosis는 특정 ligand를 선택적으로 농축하여 흡수하는 효율적 경로이다.
Selective Concentrating Mechanism
- AP2가 receptor의 cytosolic face를 선택적으로 인식 → receptor 모집
- AP2는 clathrin triskelion을 recruit하고, 이것이 다른 AP2와도 연결 → AP2가 집적되는 결과 → receptor 선택적 농축
- 따라서 clathrin-coated pit 자체가 AP2 집합체로 형성 → 특정 receptor만 농축
- 특정 ligand의 내재화 효율을 100배 이상 증가시킴
- 세포외 fluid의 minor component도 대량으로 효율적으로 흡수 가능
- 대부분의 plasma membrane 단백질은 clathrin-coated pit에 농축되지 못함
- 하나의 clathrin-coated pit이 100개 이상의 다양한 receptor를 동시에 수용 가능
LDL Receptor의 작동
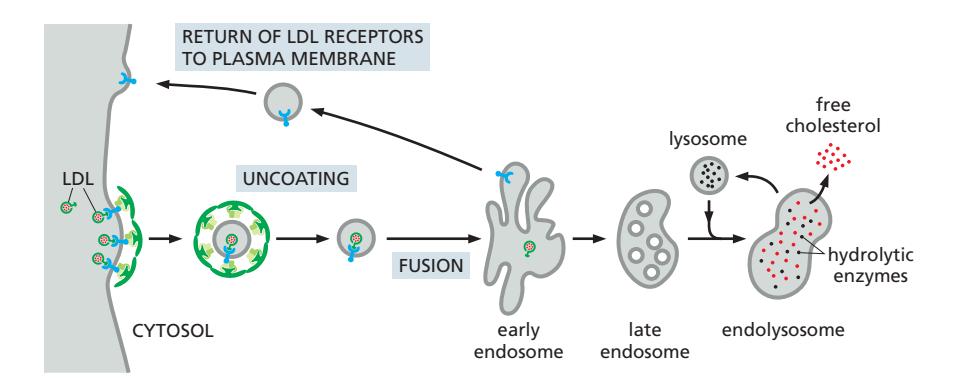
- LDL receptor: plasma membrane에 존재
- Cytoplasmic tail의 endocytosis signal → AP2 결합
- AP2가 PI(4,5)P₂에 결합하여 unlocked → cargo binding site 노출
- AP2가 clathrin triskelion 모집 → coated pit 형성
- Dynamin에 의해 pinch off → coated vesicle
- Coat shed → early endosome 융합
LDL의 처리
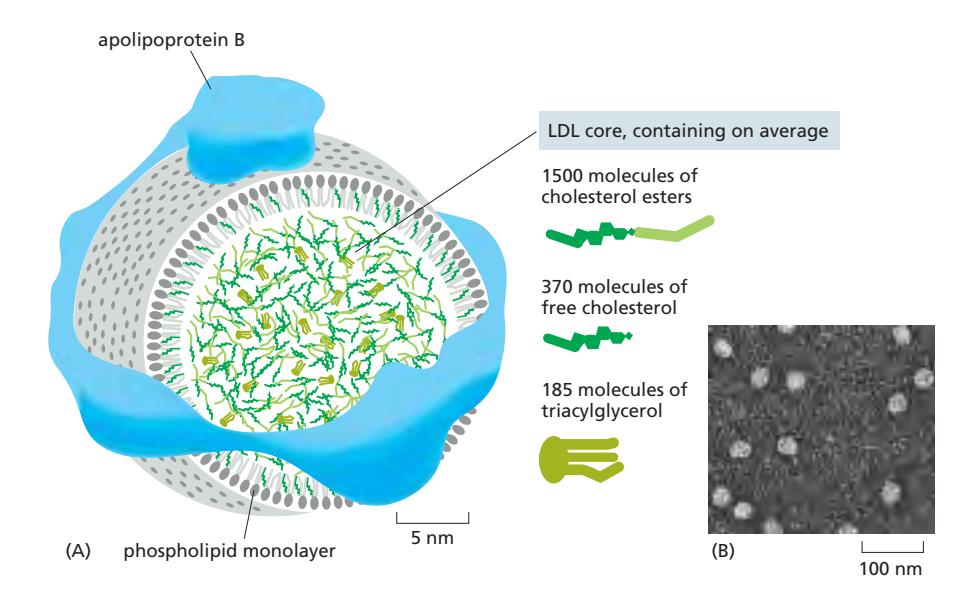
- Early endosome의 낮은 pH → LDL이 receptor에서 방출
- LDL → Late endosome → Lysosome
- Lysosome: cholesterol ester → free cholesterol (막 합성에 사용)
- LDL receptor: early endosome tubular region → plasma membrane으로 재활용
Endocytosis 이후 Receptor의 처리
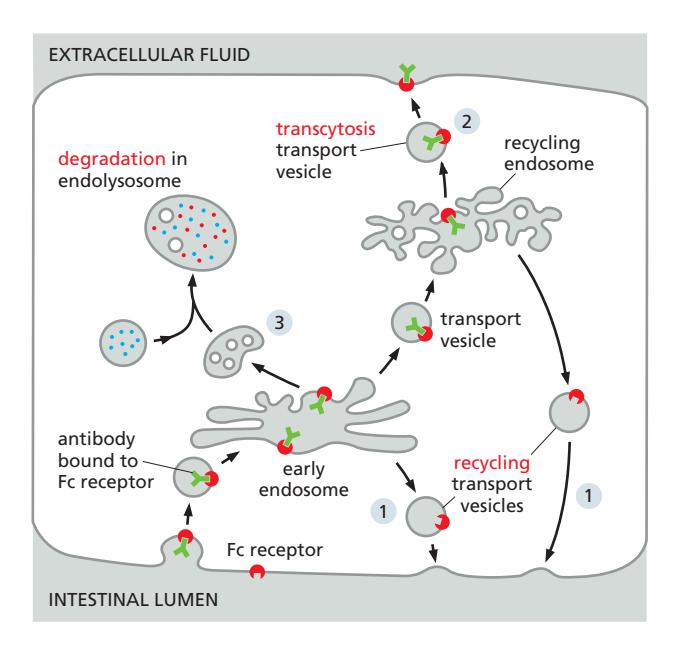
Cargo가 붙은 상태이든 아니든, receptor는 endocytosis 이후 세 가지 경로 중 하나로 처리된다:
| 경로 | 내용 |
|---|---|
| Recycling | Cargo를 처리한 뒤 receptor를 원래 plasma membrane 위치로 재활용 (예: LDL receptor, transferrin receptor) |
| Transcytosis | Cargo를 세포의 반대편 막으로 운반 (예: 장관강 → 세포외 fluid) |
| Degradation | Receptor가 endolysosome/lysosome에서 분해됨 (예: ubiquitinated signaling receptor → MVB pathway → lysosome) |
9. Exocytosis
Exocytosis는 transport vesicle이 plasma membrane과 융합하여 내용물을 세포 외부로 방출하는 과정이다.
두 가지 경로
1. Constitutive Secretory Pathway (default)
- 모든 세포에서 지속적으로 작동
- 특별한 신호 없이 자동으로 진행
- Plasma membrane 성분, ECM 단백질 전달
2. Regulated Secretory Pathway
- 특수화된 분비 세포에만 존재
- Extracellular signal에 의해 조절 → Ca²⁺가 주요 trigger
- Secretory vesicle에 cargo를 저장 → 신호 시 방출
- Hormone, neurotransmitter, digestive enzyme 등 분비
Secretory vesicle 형성과 성숙:
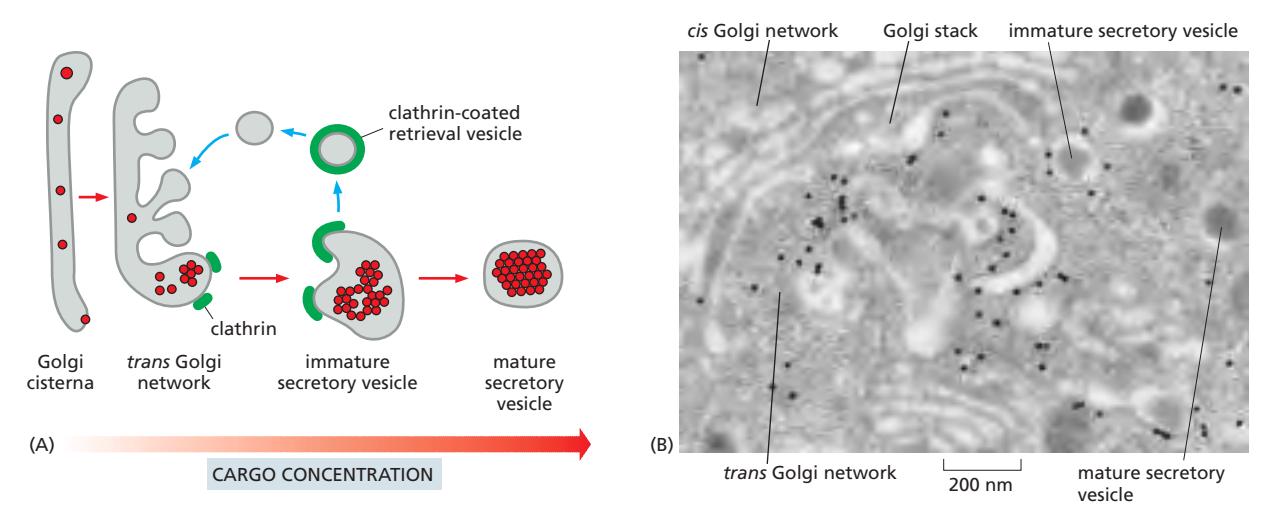
- TGN에서 aggregation: 낮은 pH와 높은 Ca²⁺ 환경이 분비 단백질 응집 촉진 (aggregation hypothesis, 정설); 또는 Chromogranins/Secretogranins 같은 granins이 sorting scaffold 역할 (sorting receptor 가설)
- Immature vesicle 형성: aggregate를 둘러싼 membrane budding; 초기에 clathrin coating 존재
- Mature secretory vesicle: clathrin 이탈, 분비 단백질 응축
Ca²⁺-triggered exocytosis:
- 신호 수신 → voltage-gated Ca²⁺ channel 개방
- Ca²⁺ influx → cytosolic Ca²⁺ 농도 급증
- Ca²⁺가 synaptotagmin에 결합 → SNARE complex 활성화 → membrane fusion
Regulated Exocytosis가 Plasma Membrane을 확장하는 경우
4 examples:
- 세포분열의 마지막 단계 Cytokinesis
- Phagocytosis 중중
- Membrane 손상 시 repair
- Fly embryo의 cellularization
10. Secretory Protein의 Proteolytic Processing
Proteolytic processing은 많은 분비 단백질이 비활성 전구체(precursor)로 합성되어 활성화되는 과정이다.
 Processing의 생물학적 이유:
Processing의 생물학적 이유:
- 보호 기능: 세포 내부에서 premature activation 방지 → 세포 손상 방지 (예: digestive enzyme은 secretory vesicle/extracellular space에서만 활성) mature form is harmful inside
- 크기 제한 극복: Enkephalin(5 aa) 같은 짧은 neuropeptide는 co-translational ER entry에 필요한 최소 길이에 미달 → polyprotein으로 합성 후 cleavage mature form is too small
- Tissue-specific enzyme 조절: 같은 precursor라도 세포 유형별 다른 processing enzyme → 다른 최종 산물
- 다양성 생성: 하나의 유전자에서 다양한 polypeptide 생성
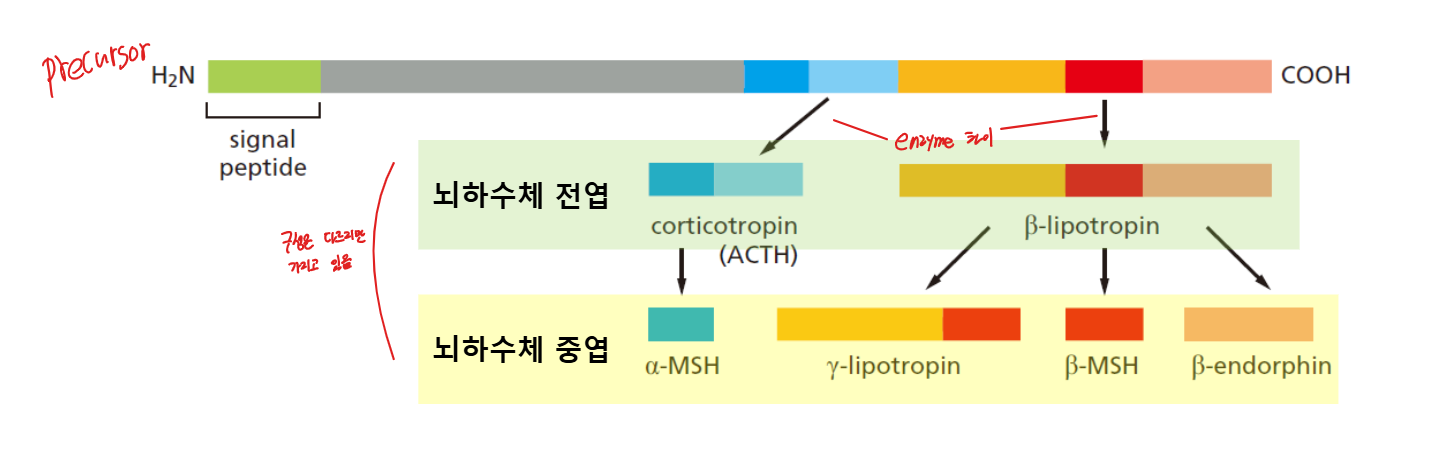
Polyprotein 예시: Proopiomelanocortin (POMC)
동일한 precursor가 서로 다른 세포에서 다른 enzyme에 의해 다른 최종 산물 생성:
- 뇌하수체 전엽 (Anterior pituitary): Corticotropin (ACTH), β-lipotropin
- 뇌하수체 중엽 (Intermediate pituitary): α-MSH, γ-lipotropin, β-MSH, β-endorphin
11. Synaptic Vesicle
Synaptic vesicle은 신경세포에서 neurotransmitter를 저장하고 millisecond 단위로 방출하는 특수화된 secretory vesicle이다.
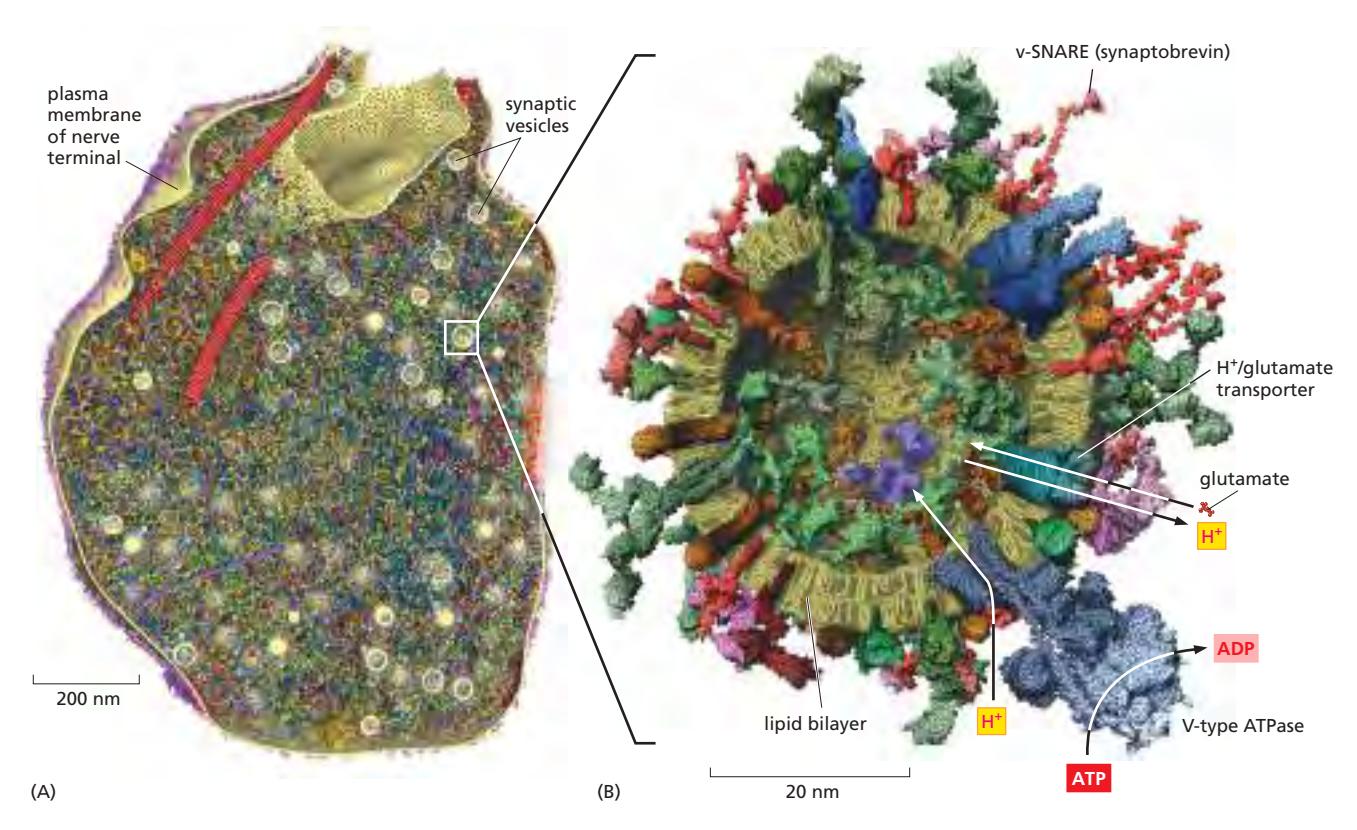
- 직경: 약 50 nm==
형성: Local Recycling
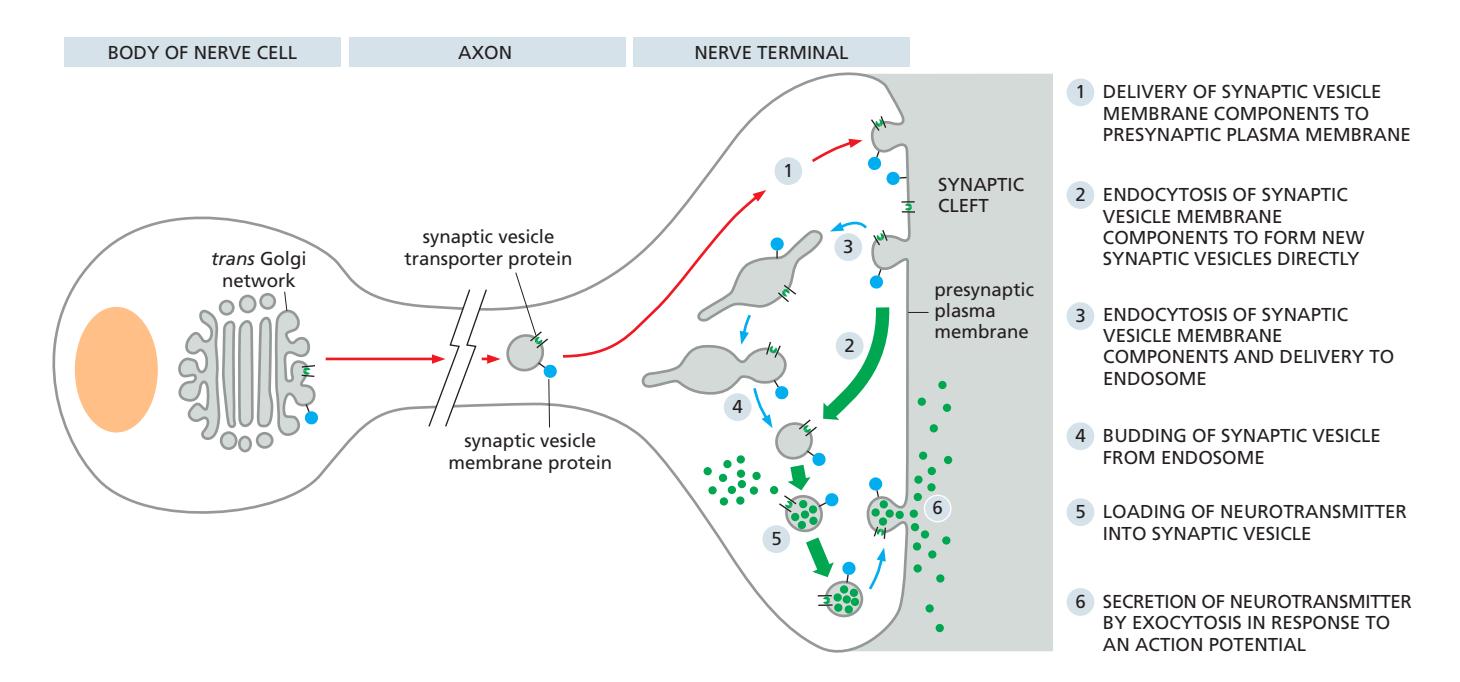
일반적인 secretory vesicle과 달리 local recycling을 통해 형성됨 즉, cytosol의 작은 neurotransmitter를 antiport를 통해 local에서 공급. ER에서 넣어져서 오는 것이 아님.
속도의 중요성:
- 신경세포는 초당 1000회 이상 발화 가능
- 빠른 vesicle 재생성 필요 → local recycling으로 해결 2 - 5 - 6 경로가 가장 빠르고 흔함 3 - 4 - 5 - 6 경로가 느리고 보조적인 경로
Priming 메커니즘
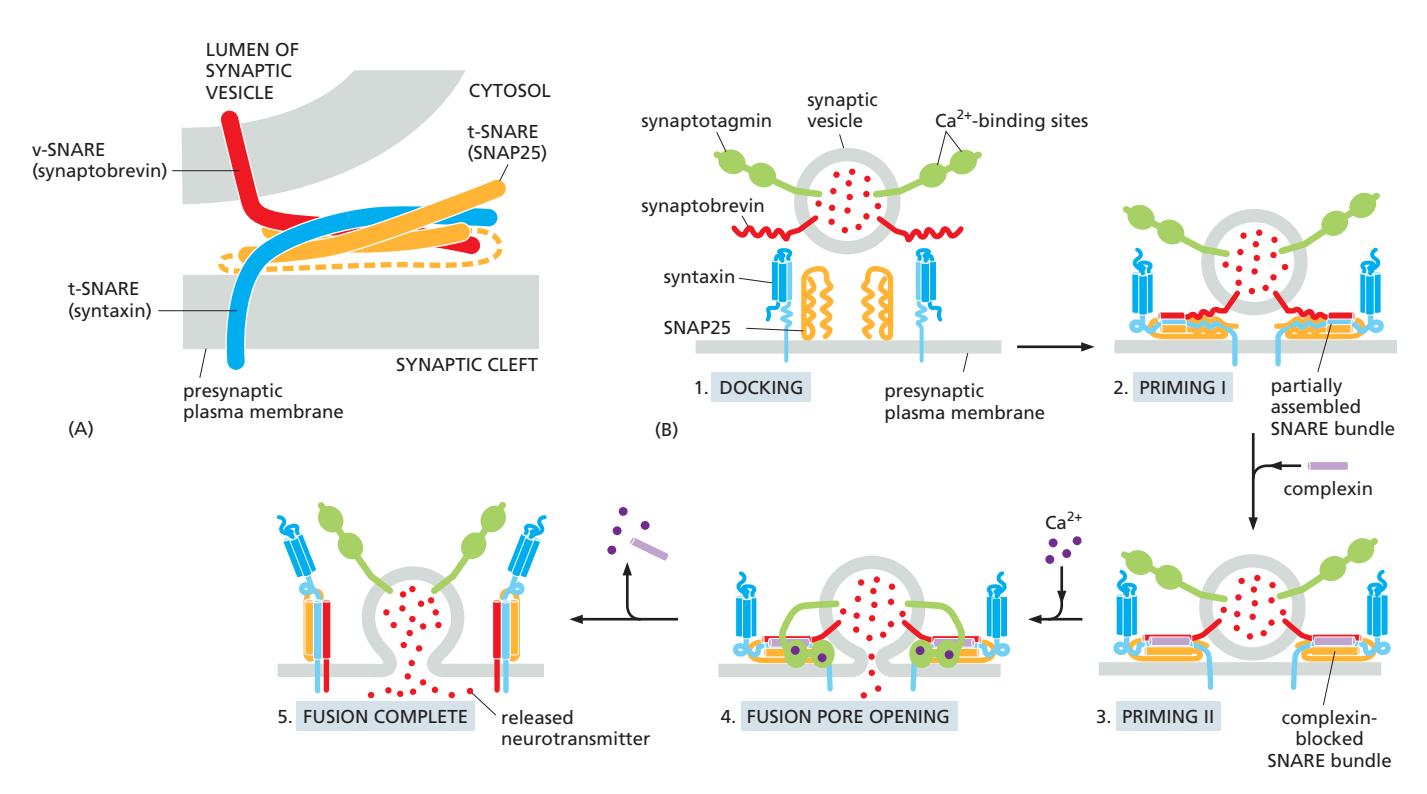
- Vesicle docking: Rab protein + tethering protein으로 presynaptic membrane에 결합
- SNARE partial assembly (Priming I): synaptobrevin + syntaxin + SNAP25가 부분적으로 assemble
- Complexin binding (Priming II): SNARE complex에 결합하여 complete zippering 방지 → metastable state 유지
- Ca²⁺ influx: synaptotagmin 활성화 → complexin displacement → SNARE fully zipper → rapid fusion
12. Polarized Cells에서의 Sorting
극성 세포(예: 상피세포)는 apical과 basolateral domain으로 단백질을 선택적으로 전달한다.
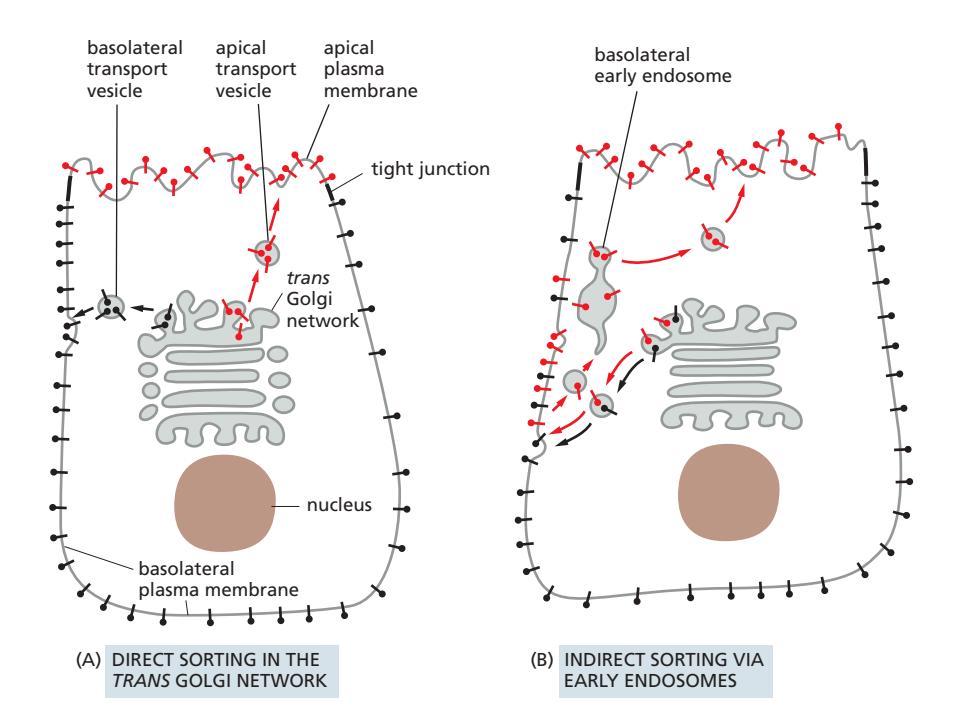
Tight Junction
- Apical domain과 basolateral domain의 경계 유지
- 단백질이 두 domain 사이를 자유롭게 이동하지 못하도록 차단
A: Direct Sorting (주요 경로)
TGN에서 직접 각 domain으로 분류:
B: Indirect Sorting (Transcytosis)
일부 단백질이 먼저 한 domain으로 전달된 후 endocytosis로 회수되어 반대 domain으로 재전달:
- 예: Hepatocyte에서 일부 apical protein이 먼저 basolateral로 → endocytosis → apical domain으로 재전달
- Signal이 두 단계 모두에서 작용
13. 전체 Intracellular Traffic 통합 흐름
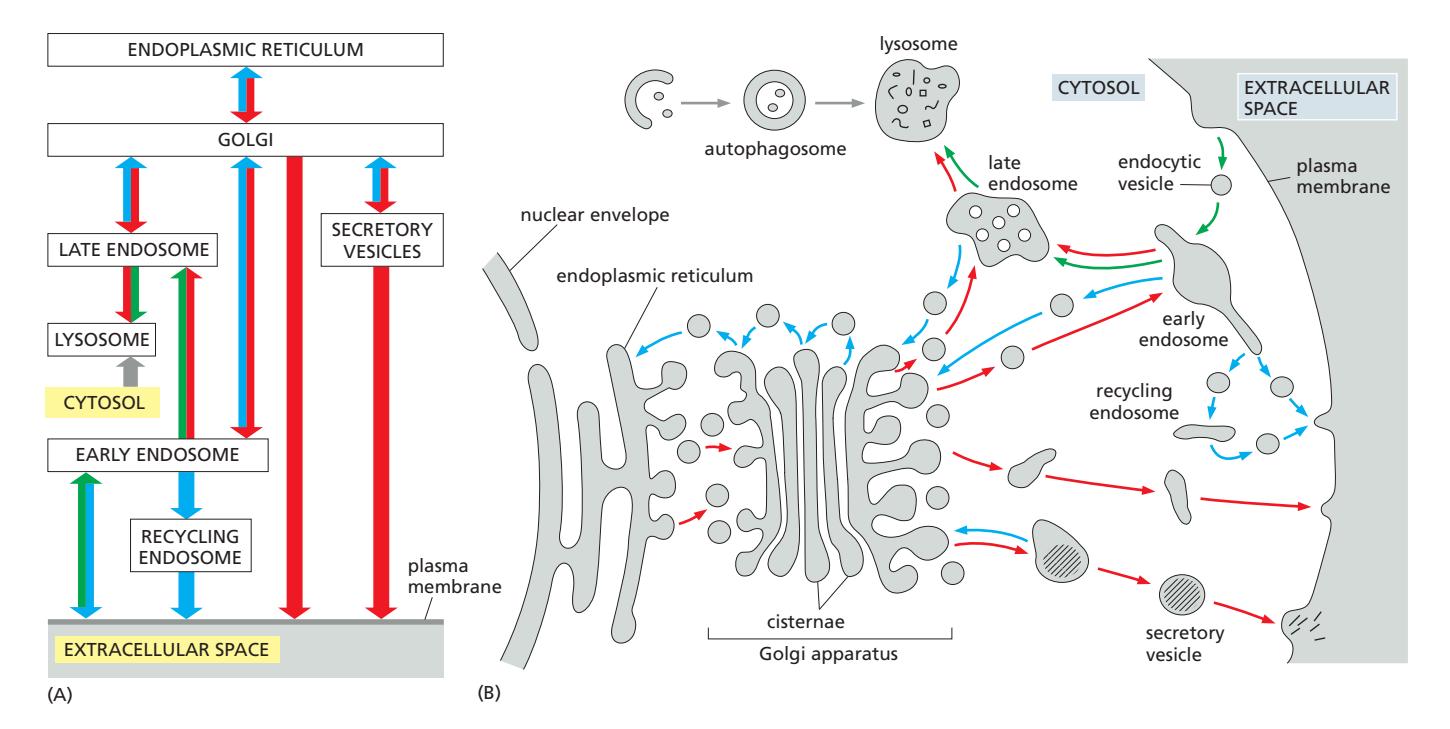
TGN
↙ ↘
M6P receptor Constitutive/Regulated secretion
↓
Early endosome (Rab5, pH ~6.0)
↙ ↘
Recycling Late endosome (Rab7, pH ~5.5)
to PM ↓ (MVB/ESCRT)
Intralumenal vesicle 형성
↓
Endolysosome / Lysosome (pH 4.5–5.0)
관련 내용 노트
- Lysosome 구조와 기능
- Lysosomal hydrolase
- M6P receptor system
- Endosome maturation
- ESCRT protein complex
- Rab5→Rab7 cascade
- Endocytosis 개요
- Receptor-mediated endocytosis (LDL)
- Caveolar endocytosis
- Macropinocytosis
- Exocytosis 개요
- Constitutive vs. regulated secretory pathway
- Synaptic vesicles
- Secretory protein proteolytic processing
- TGN에서의 direct sorting
- SNARE protein