Chapter 13-1: Intracellular Membrane Traffic 정리본
5주차 강의노트 기반 | 출처: chapter 13 강의 I 2026.pdf
1. Intracellular Membrane Traffic 개요
Intracellular membrane traffic은 세포 내 막으로 둘러싸인 구획들 사이에서 일어나는 물질 이동 체계이다. 크게 세 방향으로 나뉜다:
- Secretory (biosynthetic) pathway: ER → Golgi → plasma membrane / 분비 / lysosome
- Endocytic pathway: Plasma membrane → endosome → lysosome
- Retrieval pathway: 잘못 운반된 단백질을 원래 compartment로 되돌리는 경로
- Autophagy: 세포 자신의 성분을 lysosome으로 분해
모든 경로는 transport vesicle을 통해 이루어진다. Vesicle은 donor membrane에서 budding하여 target membrane과 fusion한다.
2. Vesicle Transport의 3단계
Vesicle transport는 다음 세 단계로 구성된다:
- Budding (출아): Coat protein이 donor membrane에서 vesicle을 형성
- Transport: Vesicle이 cytoskeleton을 따라 target compartment로 이동
- Fusion (융합): Vesicle이 target membrane과 융합하여 cargo 전달
Coated Vesicle의 종류
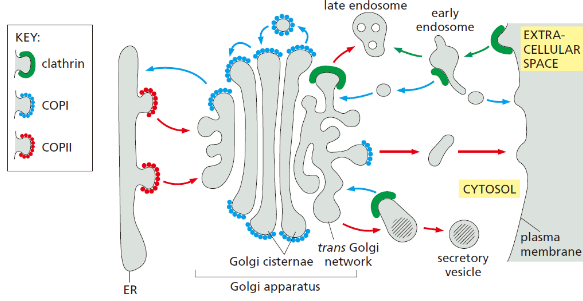
| Coat | 위치 | 방향 |
|---|---|---|
| Clathrin | Plasma membrane, TGN | Endocytosis(PI(4,5)P2가 시작) TGN→endosome (monomeric GTPase ARF가 시작.AP1과 상호작용) early endosome ,immature secretory vesicle → Golgi retrival. |
| COPI | Golgi | Retrograde (Golgi→ER) |
| COPII | ER | Anterograde (ER→Golgi) |
3. Clathrin Coat
Clathrin은 가장 잘 알려진 coat protein이다.
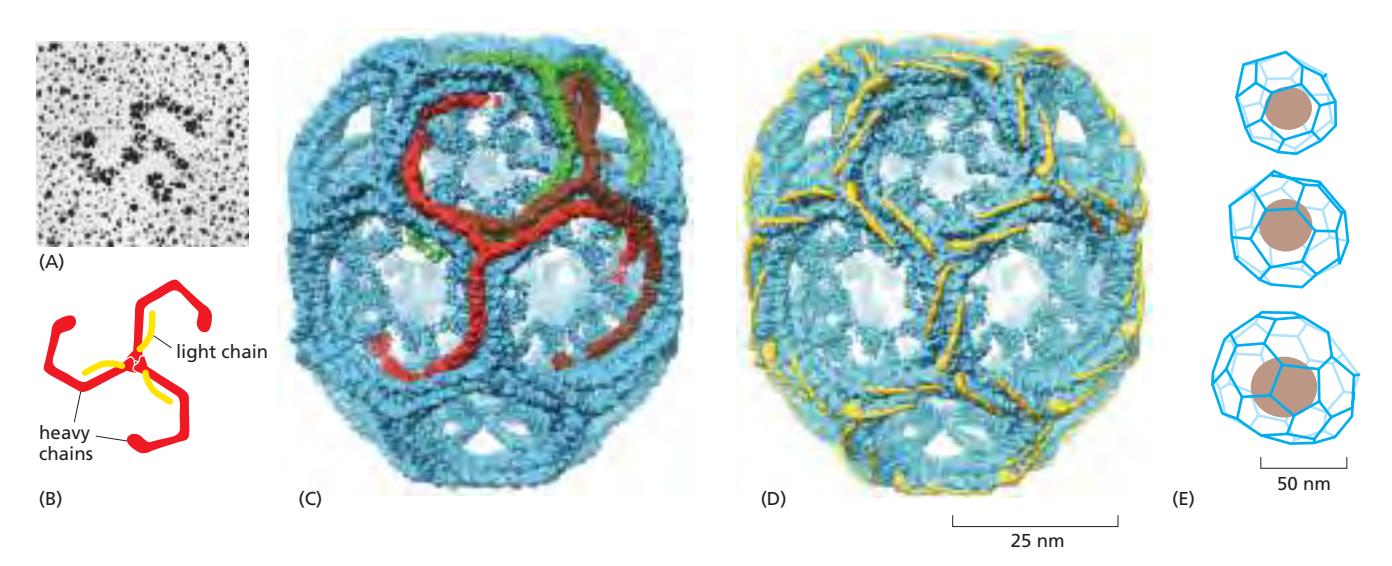
구조: Triskelion
- Triskelion: 3개의 heavy chain + 3개의 light chain으로 구성
- Heavy chain은 세 방향으로 뻗은 삼각형 구조
- 여러 triskelion이 모여 hexagon과 pentagon으로 구성된 polyhedral cage 형성 (축구공 모양)
조립과 분해 (Assembly/Disassembly)
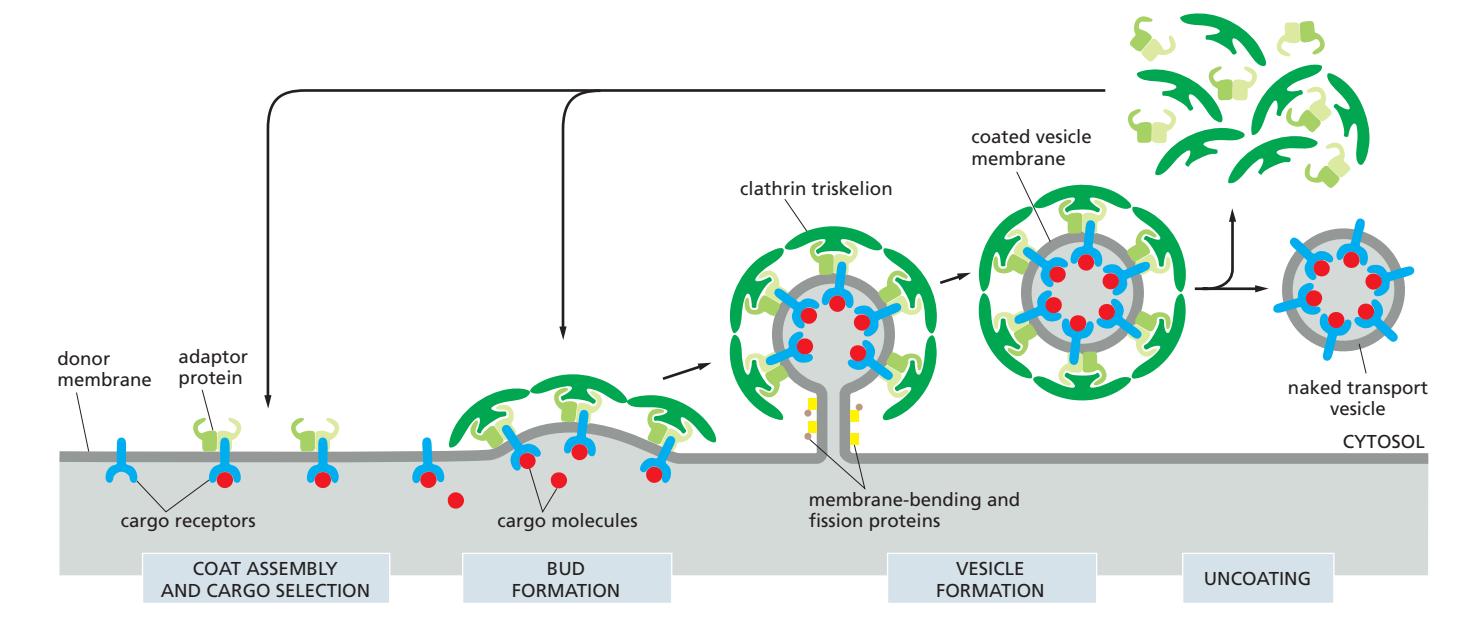
Coat assembly:
- AP2 adaptor protein이 membrane에 결합
- Adaptor가 cargo receptor와 상호작용
- Clathrin triskelion이 recruit되어 coat 형성
- Membrane curvature 증가하며 coated pit 형성
- Membrane-bending protein과 fission protein (dynamin) recruit → pinch off
Coat disassembly:
- Vesicle이 budding된 직후 발생 (목적지 도달 후가 아님)
- hsp70 chaperone이 ATP hydrolysis 에너지로 clathrin coat를 물리적으로 제거
- Phosphoinositide phosphatase가 PI(4,5)P₂를 분해하여 adaptor protein의 막 결합 약화
4. AP2 어댑터 단백질
AP2는 clathrin coat 형성의 핵심 어댑터 단백질이다.
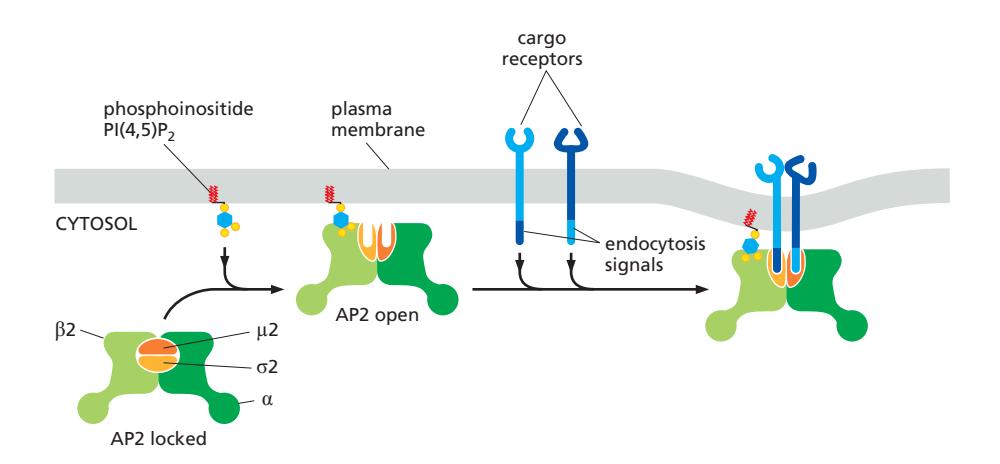
활성화 메커니즘
- 초기 상태 (locked): Cargo binding site가 차단된 비활성 구조
- PI(4,5)P₂에 결합 → conformational change → cargo binding site 노출
- Cargo motif 인식:
YXXφ (tyrosine-based)dileucine motif ([DE]XXXL[LI])
5. Phosphoinositides (PIPs): Organelle Identity Marker
Phosphoinositides (PIPs)는 막 지질의 소수 성분(<10%)이지만 organelle identity를 결정하는 핵심 marker이다.
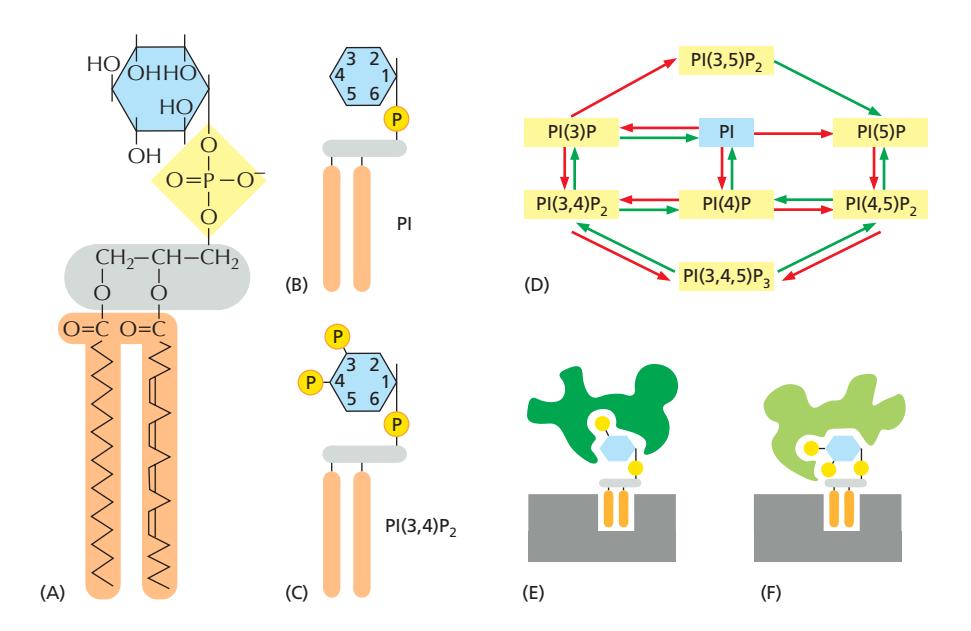
구조
- PI (phosphatidylinositol): inositol sugar head group 보유
- Inositol ring의 3’, 4’, 5’ position의 hydroxyl group이 인산화되어 다양한 PIP 생성
AP와의 관계
PIP 종류에 따라 가지고오는 Adaptor Protein이 다르다. AP가 다르면 다른 receptor를 불러온다. 결국 PIP의 조절로 ligand의 선별적 취합이 가능하다.
Compartment별 분포
| Compartment | 주요 PIP |
|---|---|
| Plasma membrane | PI(4,5)P₂ |
| Early endosome | PI(3)P |
| Late endosome | PI(3)P + PI(3,5)P₂ |
| Golgi | PI(4)P |
| ER | PI |
- 모두 cytosolic leaflet에 위치
- 각 organelle에 특이적인 kinase와 phosphatase의 분포가 PIP 특이성 결정
- PIP-binding domain을 가진 단백질이 특정 organelle로 선택적으로 recruit됨
6. Membrane-bending Proteins
Membrane-bending protein은 vesicle budding 시 막을 구부리는 역할을 한다. vesicle 형성에 꼭 필요하다.
BAR Domain Dimers
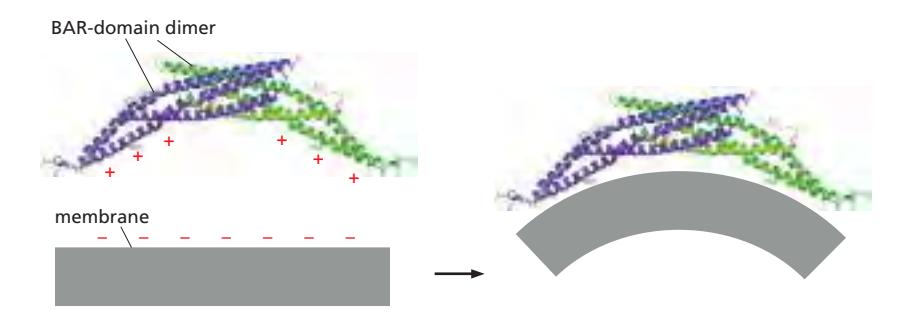
- 두 개의 subunit이 crescent-shaped dimer 형성
- Positively charged inner surface가 negatively charged lipid head group과 상호작용 → membrane을 구부림
- 굽어진 내면이 tube 형태의 membrane에 결합
- 결합 자체가 membrane에 curvature 부여
- 이미 구부러진 막에 preferential하게 결합하여 곡률 증폭
- 일부 BAR-domain protein: PIP-binding domain 보유 → 특정 membrane으로 targeting
7. Dynamin: Vesicle Fission
Dynamin은 budding vesicle을 donor membrane으로부터 분리(pinch off)하는 GTPase이다.
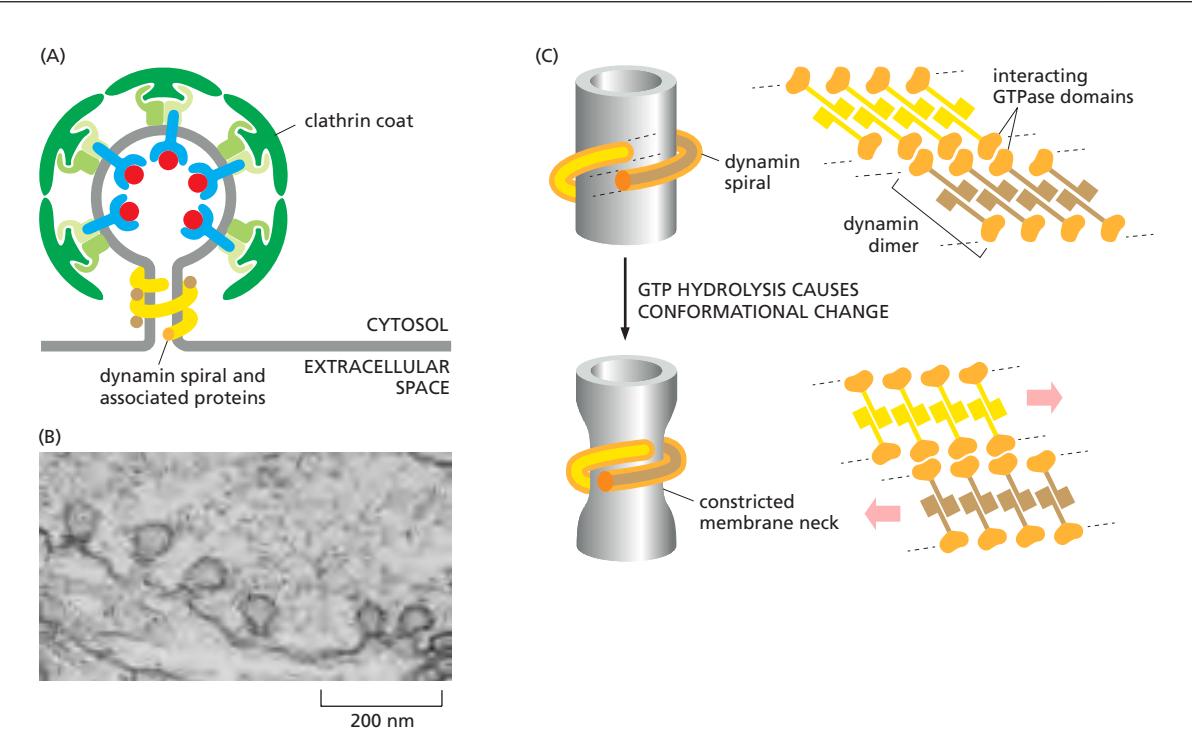
구조
- GTPase domain: GTP hydrolysis 조절
- PH domain (PI(4,5)P₂-binding domain): 막의 PI(4,5)P₂에 결합
- Middle domain/stalk: oligomerization 담당
작동 메커니즘
Assembly:
- Dynamin의 PH domain이 PI(4,5)P₂에 결합하여 막에 붙음. vesicle neck 주위에 ring 형성
- Ring이 spiral structure로 조립 (polymer 형성)
Constriction:
- GTPase domain이 dimerize하면서 Bound GTP hydrolysis
- Dynamin의 conformational change
- Dynamin ring constriction → underlying membrane neck constrict
- noncytosolic leaflet이 모임
- Fission 완성
Dynamin dimer: G-domain이 adjacent rung의 G-domain과 dimerize → GTPase 활성화 → conformational change → constriction
8. Monomeric GTPases: Coat Assembly 조절
Coat-recruitment GTPase는 특정 compartment에서만 coat assembly를 활성화하는 분자 스위치이다.
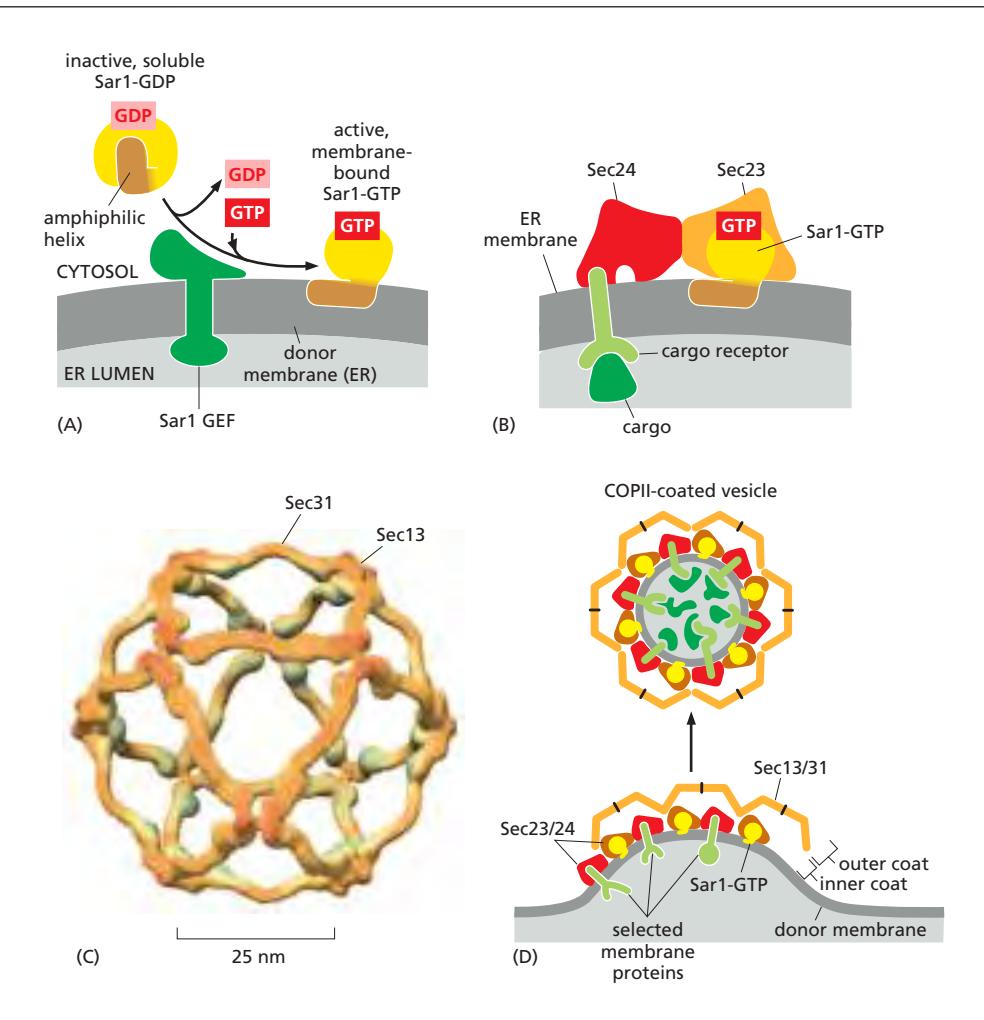
종류와 역할
| GTPase | 작동 위치 | 조절하는 coat |
|---|---|---|
| Sar1 | ER membrane | COPII coat |
| ARF proteins | Golgi membrane | COPI coat, Clathrin coat. |
| ARF는 AP1을 활성화한다. 이 역시 clathrin coat를 형성한다. AP2와 AP1은 다른 adaptor로, 다른 receptor에 특이성을 보인다. |
GDP/GTP Cycle
- GEF (Guanine nucleotide exchange factor): GDP → GTP 교환 촉매 → 활성화
- GAP (GTPase-activating protein): GTP hydrolysis 촉진 → 불활성화
Sar1에 의한 COPII Coat Assembly
- ER membrane embedded Sar1 GEF(Sec12)가 세포질의 Sar1-GDP 활성화
- GEF가 GDP를 방출 → 세포질의 GTP가 결합 → Sar1-GTP 형성
- Sar1의 N-terminal amphiphilic helix 노출 → ER membrane cytoplasmic leaflet에 wedge처럼 삽입
- Sar1-GTP가 Sec23/Sec24 (inner coat, adaptor) 모집
- Sec23/Sec24 위에 Sec13/Sec31 (outer coat) 조립
- Vesicle budding 완성
ARF도 같은 원리: Golgi membrane의 ARF GEF에 의해 활성화 → lipid anchor로 Golgi membrane에 삽입 → COPI/clathrin coat 모집
Not all transport vesicles are spherical
procollagen이 vesicle에 담길 때도 COPII coat가 vesicle을 만든다. 그런데 긴 vesicle에 담기며 ER에서 budding한다.
9. Rab Proteins: 수송 정확도의 핵심
Rab GTPase는 vesicle의 tethering, docking, fusion의 정확도를 담당한다.
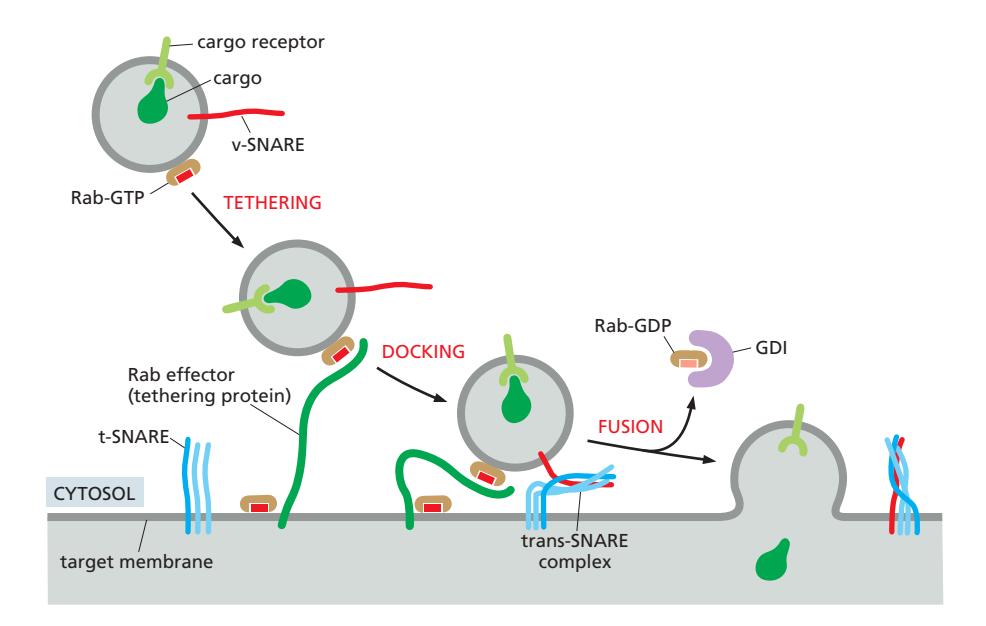
Rab의 작동 원리
Vesicle과 target membrane에 동일한 Rab GTPase가 있다. vesicle의 Rab은 Rab effector에 의해 tethering 되어 t-SNARE가 있는 곳까지 recruit된다. 결국 fusion이 일어남남
| 상태 | 결합 분자 | 위치 |
|---|---|---|
| GTP-bound (활성) | Rab effectors | Target membrane |
| GDP-bound (비활성) | GDI | 세포질 |
Rab cycle:
- **GDI (GDP Dissociation Inhibitor)**가 GDP-Rab을 세포질에 유지
- Membrane의 GEF가 GDI로부터 Rab을 떼어내고 GTP와 교환
- GTP-Rab이 membrane에 결합 및 effector 모집
- Effector: tethering factor, motor protein, SNARE 조절인자 등
- GTP 가수분해 후 GDI가 GDP-Rab을 다시 세포질로 추출
Rab5 Domain 형성
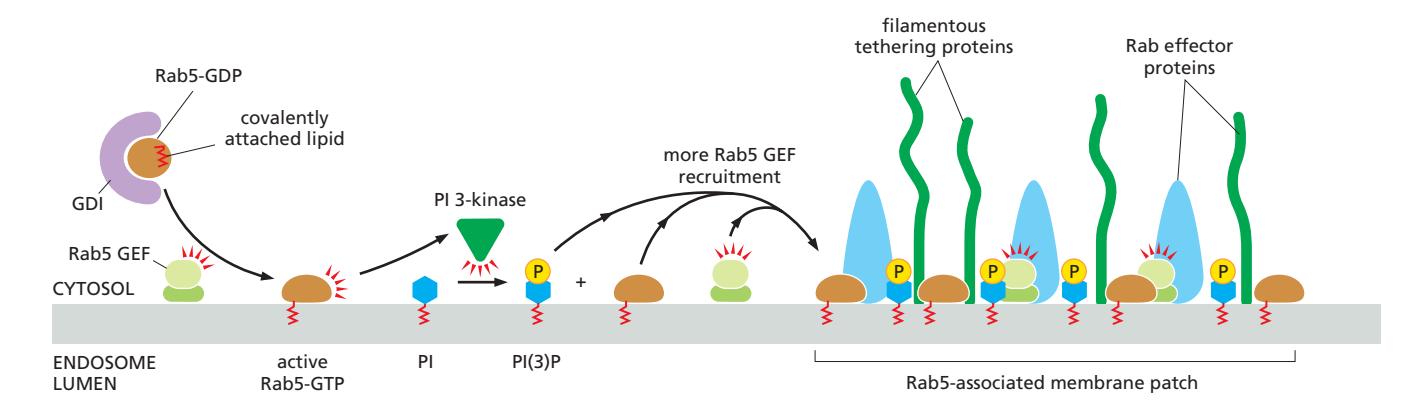
- Rab5가 **PI3K (PI 3-kinase)**를 effector로 모집
- PI3K가 PI(3)P 생성 → PI(3)P-binding effectors 추가 모집
- Self-amplifying loop: Rab5-GTP가 더 많은 Rab5-GTP를 모집
- PI(3)P enrichment가 early endosome의 identity 정의
Vesicle Tethering과 Docking
Long-range tethering:
- Rab effector (filamentous protein)가 target membrane에서 vesicle을 먼 거리(200 nm)에서 포착
- Vesicle의 Rab-GTP와 target membrane의 Rab effector가 상호작용
- 이때 vesicle의 Rab과 target membrane의 Rab은 같은 종류여야 함
SNARE-mediated docking:
- v-SNARE와 t-SNARE pairing 촉진 → membrane fusion 유도
Rab inactivation:
- Fusion 후 Rab GAP가 Rab-GTP를 가수분해 → Rab-GDP
- GDI가 Rab-GDP를 membrane에서 추출 → cytosol로 복귀
Rab5→Rab7 Cascade
- Rab5-associated membrane patch가 시간이 지나면서 Rab7-associated patch로 교체
- Rab5가 recruit 하는 effector 중에 Rab7 GEF가 있어 Rab7을 recruit
- Rab7가 recruit 하는 protein 중에 Rab5 GAP가 있어 Rab5 domain을 inhibit.
- 이 전환이 일방향적이고 비가역적: early endosome → late endosome 성숙
- Rab5→Rab7 전환 실패 시 cargo가 lysosome에 도달 불가
10. SNARE Proteins: Membrane Fusion
SNARE protein은 vesicle과 target membrane의 융합을 직접 촉매한다.
분류
- v-SNARE (vesicle-SNARE): transport vesicle membrane에 위치. Single polypeptide
- 대표 예: Synaptobrevin
- t-SNARE (target-SNARE): target membrane에 위치. 보통 3개 protein으로 구성
- 대표 예: Syntaxin (1 helix) + SNAP25 (2 helices)
Fusion 5단계
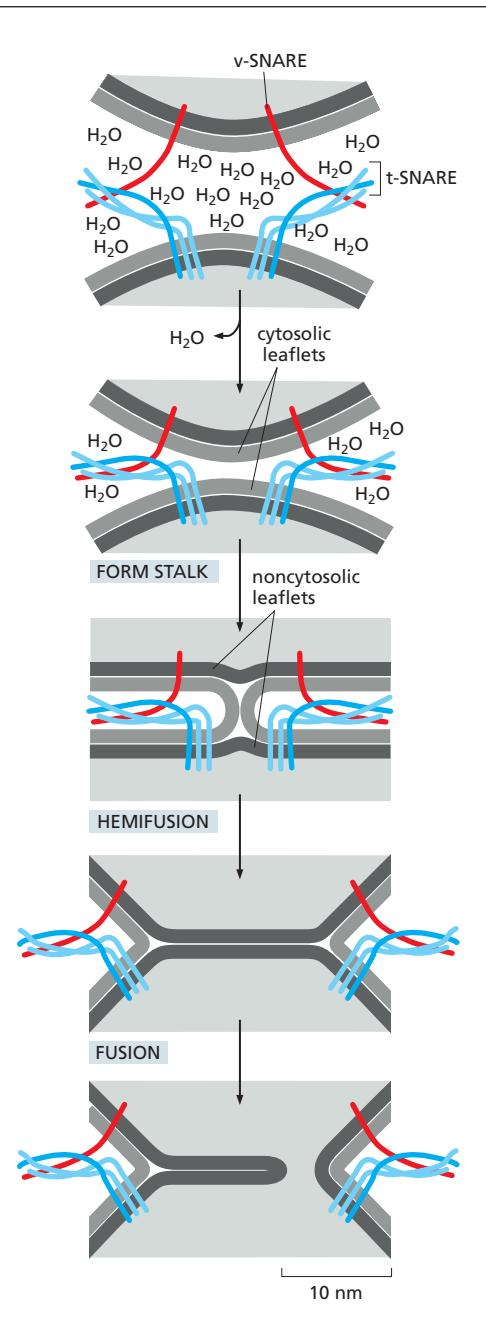
- Membrane Apposition: 두 membrane이 tight하게 lock
- Expels water molecules
- SNARE pairing의 stability가 energy 제공
- Stalk Formation: cytosolic leaflet끼리
- Hemifusion:
- noncytosolic leaflet끼리 New bilayer 형성 (hemifusion 또는 half-fusion)
- New bilayer rupture:
- Complete fusion reaction
NSF
NSF는 사용된 v-snare와 t-snare의 꼬임을 풀어주는 단백질복합체. accessory protein과 함께 작용함
AAA ATPase=unfoldase=NSF
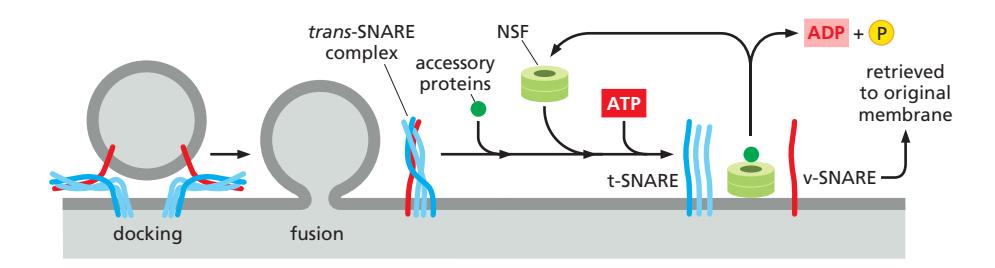
SNARE-mediated fusion의 에너지는 궁극적으로 NSF의 ATP hydrolysis에서 유래. NSF가 SNARE를 분리하는 데 ATP 소비 → 분리된 SNARE가 다시 assembly하며 fusion을 drive하는 cycle.
Homotypic vs Heterotypic Fusion
- Heterotypic fusion (대부분의 경우): 서로 다른 compartment의 membrane이 융합. v-SNARE + t-SNARE pairing
- Homotypic fusion: 같은 종류의 membrane끼리 융합. 양쪽 membrane에 v-SNARE와 t-SNARE 모두 존재
- 예: ER-derived vesicle끼리 융합하여 VTC(vesicular tubular clusters) 형성, endosome끼리 융합
Synaptic Vesicle의 특수한 SNARE
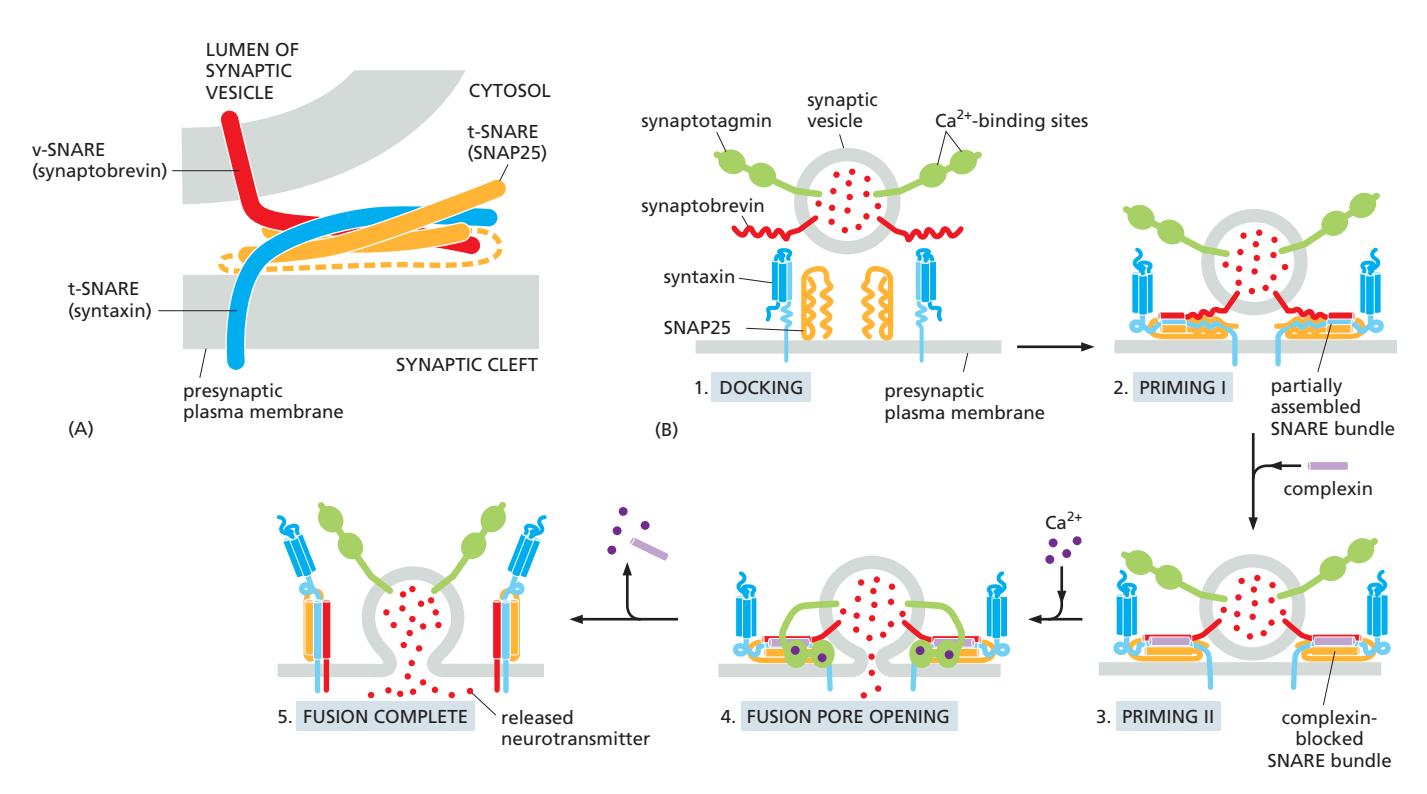 Component:
Component:
- v-SNARE: Synaptobrevin (transmembrane)
- t-SNARE: Syntaxin (transmembrane, 1 helix) + SNAP25 (peripheral, 2 helices)
조절 메커니즘 (Complexin + Synaptotagmin):
- SNARE partially assemble → primed state
- Complexin이 premature fusion 방지 (metastable state로 freeze)
- Ca²⁺ influx → Synaptotagmin (Ca²⁺ sensor) 활성화
- Complexin release
- SNARE fully zipper → rapid fusion
- Neurotransmitter release (millisecond scale)
11. ER-to-Golgi Transport
ER에서 Golgi로의 수송은 COPII vesicle을 통해 이루어진다.
12. Vesicular Tubular Cluster (VTC)
VTC는 ER에서 Golgi로 이동하는 중간 수송 구획이다.
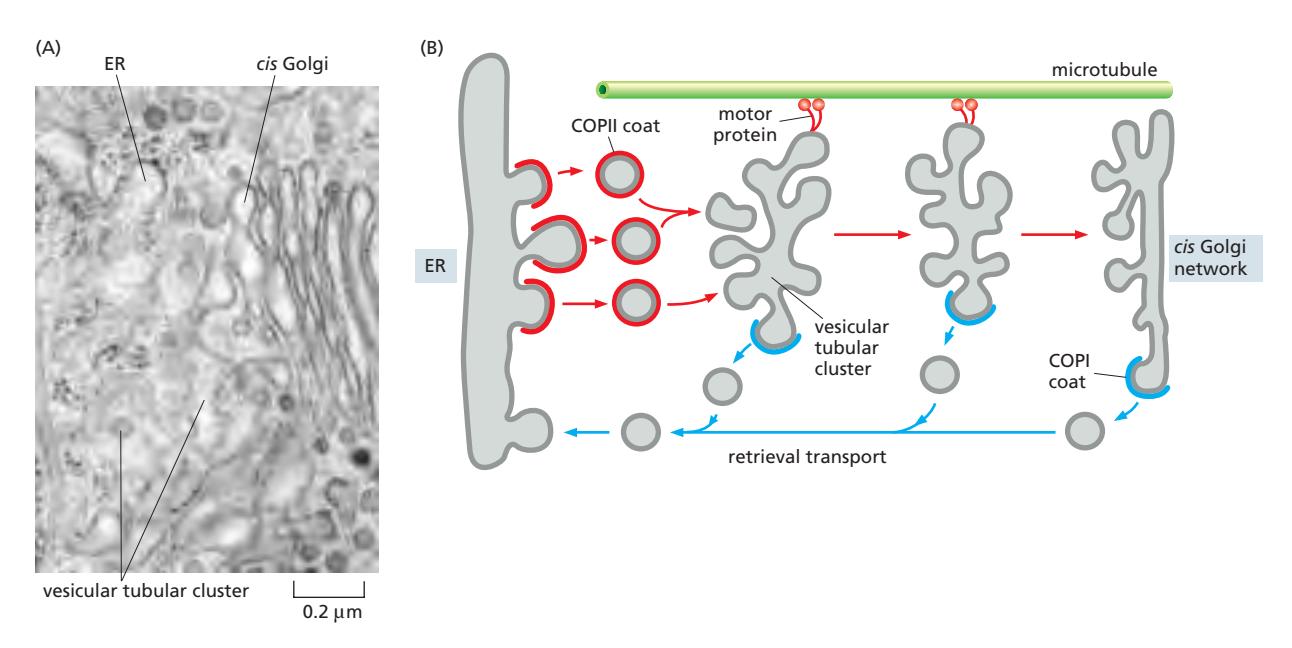
형성
- COPII-coated vesicle이 coat를 shed한 후 homotypic fusion (SNARE mediated)
- Convoluted 모양의 vesicular tubular cluster 형성
- 지속적으로 새로 생성되어 ER → Golgi로 material 운반
- 이 운반은 microtubule을 따라 motor protein에 의해 진행됨됨
Golgi로의 이동
- Microtubule을 따라 motor protein에 의해 Golgi로 이동
- VTC가 Golgi와 fusion하여 cargo 전달
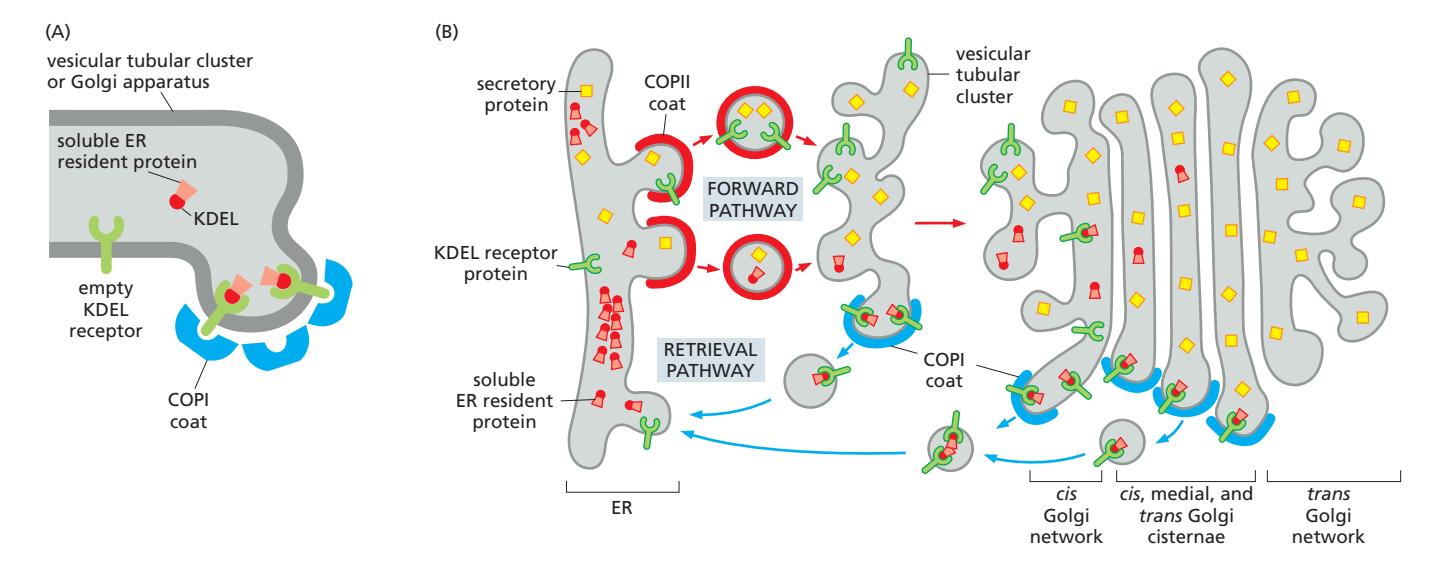
13. Retrieval Pathway: ER Resident Protein 회수
Retrieval pathway는 실수로 ER을 떠난 단백질을 되돌리는 경로이다.
COPI-coated Vesicle
- VTC에서 즉시 COPI-coated vesicle budding 시작
ER Retrieval Signal
막 단백질 신호: KKXX
- C-terminal에 two lysine + any two amino acid: KKXX sequence
- COPI coat에 직접 결합 → COPI vesicle로 ER 귀환
- ER-Golgi interface에서 기능하는 SNARE, cargo receptor에서 발견
가용성 단백질 신호: KDEL
- C-terminal의 KDEL sequence (Lys-Asp-Glu-Leu)
- BiP 등 soluble ER resident protein이 보유
- KDEL receptor (multipass transmembrane protein)가 인식 → KDEL protein은 기능 중단, COPI vesicle로 packaging
KDEL receptor cycling:
- COPII를 타고 Golgi로 이동
- Golgi (pH ~6.5–6.7): KDEL sequence에 강하게 결합 → COPI vesicle로 ER 귀환
- ER (pH ~7.2): 약한 결합 → KDEL-bearing protein 방출
- Critical histidine: Golgi의 낮은 pH에서 protonated → KDEL과의 interaction 강화
- KDEL receptor가 ER↔Golgi 사이를 shuttle하며 escaped protein 회수
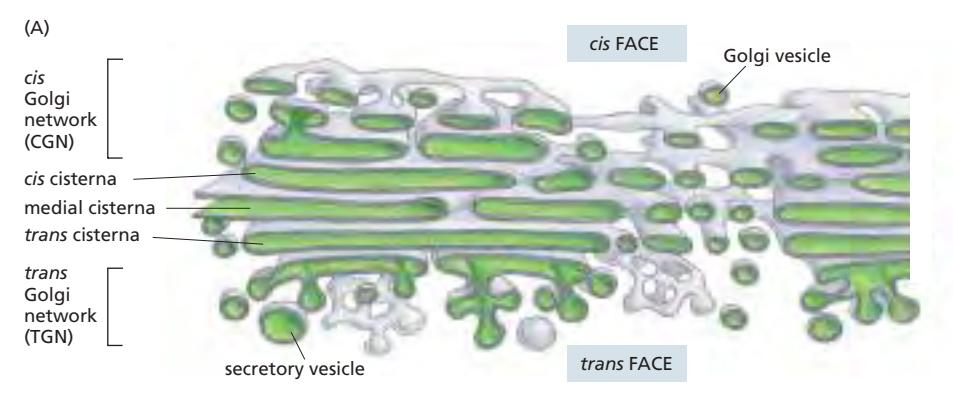
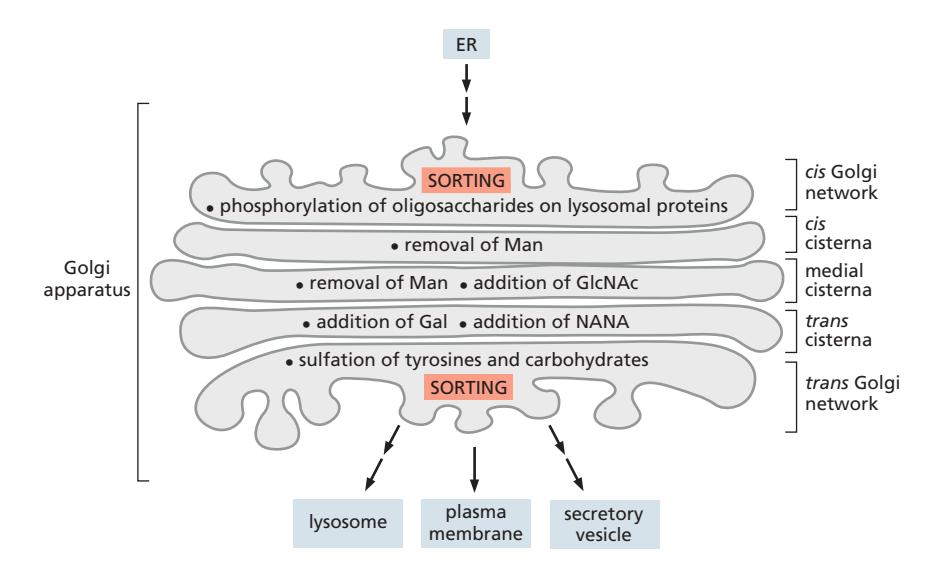
14. Golgi Apparatus
Golgi apparatus는 secretory pathway의 중심 processing 기관이다.
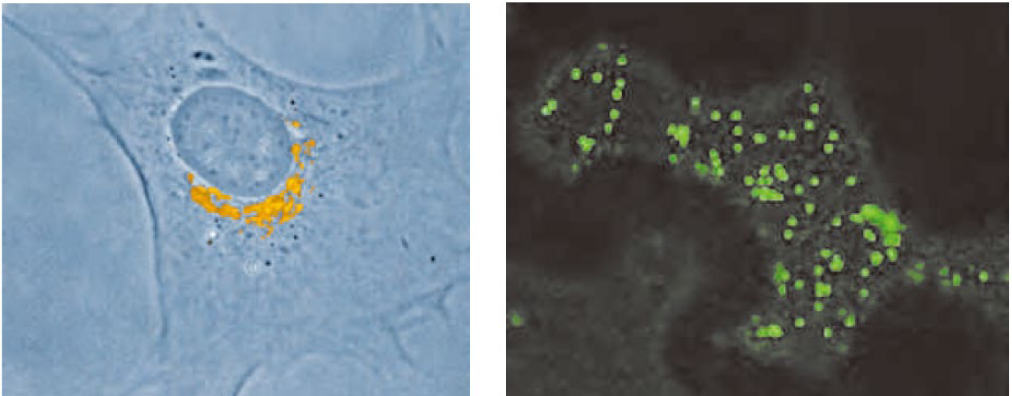 좌측이 동물세포의 골지. 핵 주위에 위치함. 우측이 식물세포의 골지. 세포 내에 산재함.
microtubule에 의한 가이드 유무 차이.
동물세포도 microtubule을 없애면 퍼지게 될 것.
좌측이 동물세포의 골지. 핵 주위에 위치함. 우측이 식물세포의 골지. 세포 내에 산재함.
microtubule에 의한 가이드 유무 차이.
동물세포도 microtubule을 없애면 퍼지게 될 것.
Cis/Medial/Trans 구획
| 구획 | 위치 | 주요 기능 |
|---|---|---|
| Cis Golgi | ER에 가장 가까운 면 | M6P tagging, 초기 processing |
| Medial cisternae | 중간 | 중간 단계 glycosylation |
| Trans Golgi | ER에서 먼 면 | 최종 glycosylation 완성 |
| TGN | Trans cisterna와 연속 | 단백질 sorting 및 발송 |
- 초기 작용 효소: 주로 cis cisternae
- 나중 작용 효소: 주로 trans cisternae
- 효소 분포는 stack 전체에 graded 방식으로 분포
N-linked Oligosaccharide Processing
 ER에서 먼저 Asn/X/Ser or Thr 서열의 Asn에 부착해서 옴.
Core region인 2GlcNAc+3Man은 항상 고정적으로 존재.
ER에서 먼저 Asn/X/Ser or Thr 서열의 Asn에 부착해서 옴.
Core region인 2GlcNAc+3Man은 항상 고정적으로 존재.
| 구획 | 가공 |
|---|---|
| Cis + Medial | Mannose 제거, N-acetylglucosamine 추가 |
| Trans + TGN | Galactose 추가, Sialic acid 추가 |
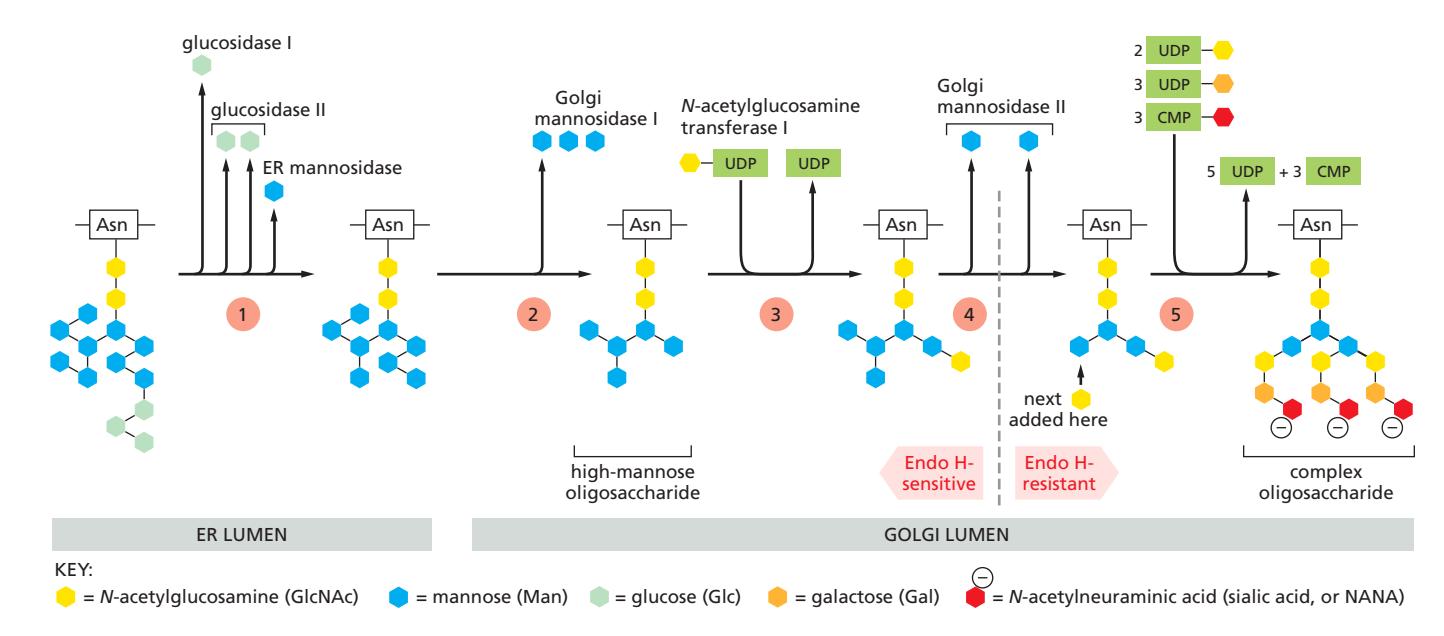
- ER에서 Glucose와 특정 Mannose를 제거하고 Golgi로 옴.
- Golgi mannosidase I가 3개의 Man 제거
- GlcNAc transferase I가 GlcNAc 첨가
- Mannosidase II가 2개의 Man제거
- GlcNAc, Gal, Sialic acid 추가
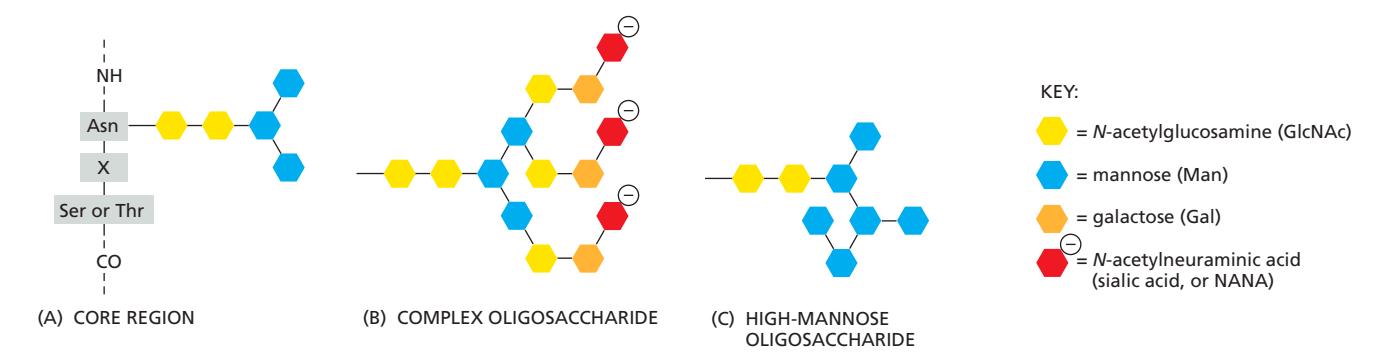 이 과정에서 Mannosidase에 노출이 쉬운, 넓은 표면의 당화는 complex form으로 되기 쉽고,
mannosidase 접근이 어려운, 단백질 내부에 묻힌 당화는 high mannose form이 되기 쉽다.
이 과정에서 Mannosidase에 노출이 쉬운, 넓은 표면의 당화는 complex form으로 되기 쉽고,
mannosidase 접근이 어려운, 단백질 내부에 묻힌 당화는 high mannose form이 되기 쉽다.
- 기질: 활성화된 sugar nucleotide (UDP-GlcNAc, UDP-Gal, CMP-NANA)
Glycosylation의 목적
protein folding promotion
- Folding이 이루어지고 있는 중간체의 용해도를 늘려 응집을 방지하는 역할
- N-연결된 올리고당의 순차적 변형은 단백질 폴딩의 진행 정도를 나타냄 (glyco-code)
proteolytic enzyme에 의한 digestion 방지
pathogen으로부터 보호, 세포-세포 recognition
세포 신호 전달의 조절 기능
Cisternal Maturation Model vs. Vesicle Transport Model
| 모델 | 특징 |
|---|---|
| Vesicle transport model | Cisternae는 고정. Cargo가 vesicle로 앞 방향 이동 COPI가 forward, backward 모두 사용됨 |
| Cisternal maturation model | Cisternae 자체가 cis→trans로 이동하며 성숙. Golgi enzyme은 COPI vesicle로 역행 이동하여 위치 유지 |
| vesicle transport model이 지지되었던 이유 | |
| 화물+/효소- 골지와 화물-/효소+골지를 섞는 실험에서 화물이 정상 처리됨. 그런데 이건 maturation model로도 설명 가능 |
Cisternal maturation model이 지지되는 이유:
- Procollagen처럼 vesicle에 포장하기 너무 큰 cargo의 이동이 관찰됨(거대 분자 관찰)
- organelle identity가 순차적으로 변화함이 실험적으로 관찰됨(실시간 현미경 찰)
요약 흐름도
ER exit site
↓ (COPII vesicle)
Vesicular Tubular Cluster (VTC)
↓ (microtubule transport)
Cis Golgi → Medial → Trans → TGN
↑
COPI retrograde vesicle (Golgi enzyme 역행, ER retrieval)
↑
KKXX / KDEL signal로 회수
관련 내용 노트
- Vesicle transportation 개요
- Clathrin coat 구조와 기능
- AP2 어댑터 단백질
- PIP — organelle identity marker
- Dynamin에 의한 vesicle fission
- ARF GTPase — coat assembly 조절
- Rab GTPase cycle
- Rab cascade — Rab5→Rab7
- SNARE protein — membrane fusion
- SNARE에 의한 vesicle fusion 메커니즘
- ER-Golgi transport
- VTC
- ER retrieval pathway
- Golgi 기능적 구획화