세포골격 (2) 정리 — Microtubules & Intermediate Filaments
8주차 강의 (Cytoskeleton 2) 전체 내용 요약. 상세 내용은 300_Microtubules_MOC, 200_Intermediate filaments_MOC 참조.
Microtubules
개요
Microtubules은 tubulin 단백질로 만들어진 길고 속이 빈 원통형 구조이다. 외경 25 nm로 actin filament보다 훨씬 단단하며(persistence length 수 mm vs. actin ~10 μm), 길고 곧은 형태를 유지한다. 세포 내에서는 한쪽 끝이 MTOC(centrosome)에 결합된 경우가 많다.
Structure of a Microtubule
→ structure of a microtubule 참조
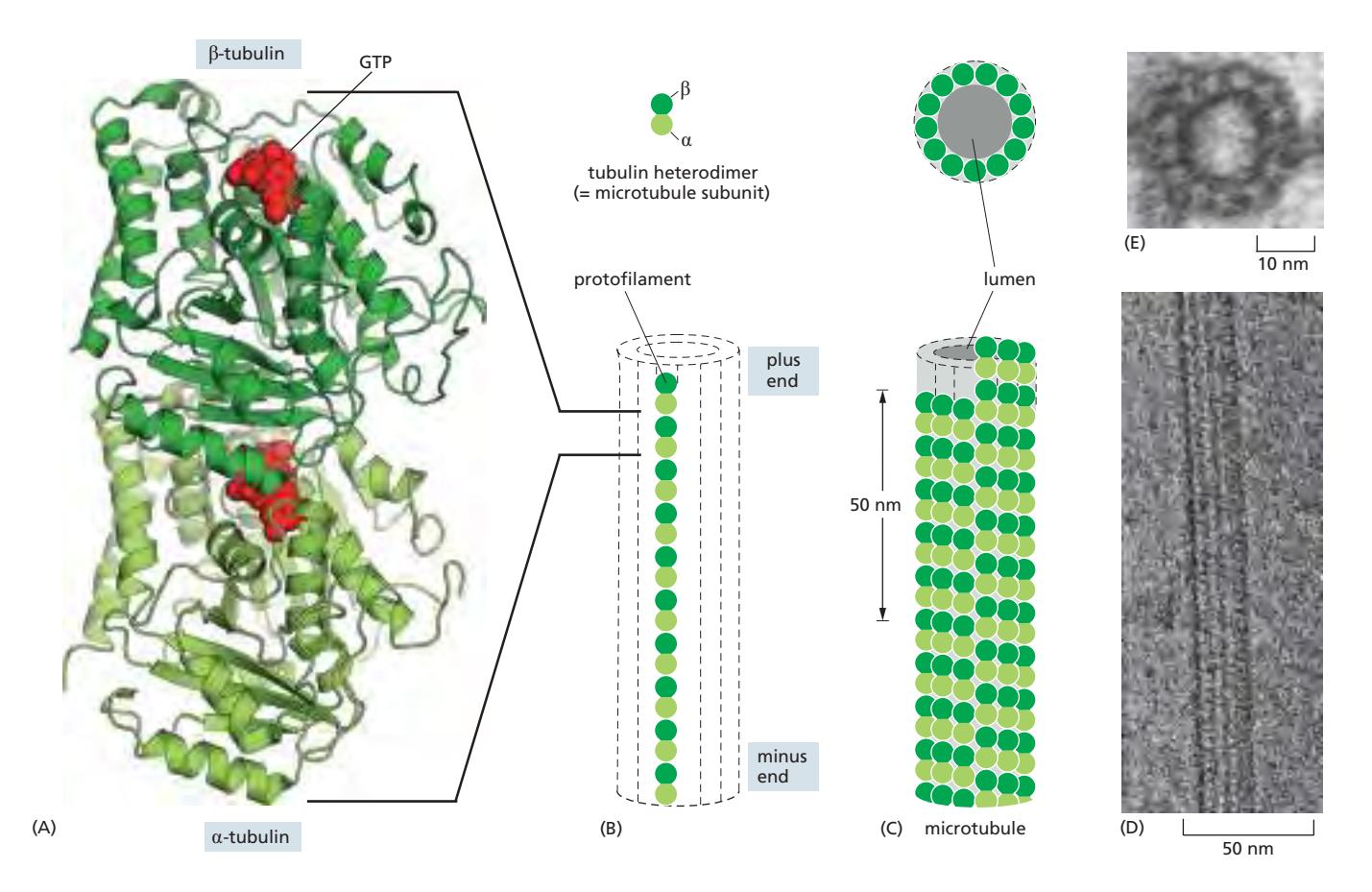 Fig 16–42. Microtubule 구조. (A) αβ-tubulin 이종이합체. (B) 13개의 protofilament가 측면으로 결합하여 hollow cylinder 형성. (C) 단일 microtubule 전자현미경.
Fig 16–42. Microtubule 구조. (A) αβ-tubulin 이종이합체. (B) 13개의 protofilament가 측면으로 결합하여 hollow cylinder 형성. (C) 단일 microtubule 전자현미경.
조립 단위: αβ-tubulin 이종이합체(heterodimer)가 중합하여 만들어진다.
- 각 tubulin 이합체의 tubulin 소단위에 GTP 결합 부위 존재
- αβ-tubulin 이합체가 protofilament를 이루며, 13개의 protofilament가 측면으로 결합하여 외경 25 nm, 내경 약 15 nm의 빈 원통 형성
- 모든 subunit이 같은 방향을 향하므로 극성(polarity) 존재:
- Plus end (β-tubulin 노출): 성장이 빠른 쪽; GTP 결합, 가수분해, 분리 모두 가능
- Minus end (α-tubulin 노출): 성장이 느린 쪽; 세포 내에서 대개 centrosome에 고정. GTP는 분해되지 않고 분리되지도 않는 구조 유지용
Preferential Growth of Microtubules
→ preferential growth of microtubules 참조
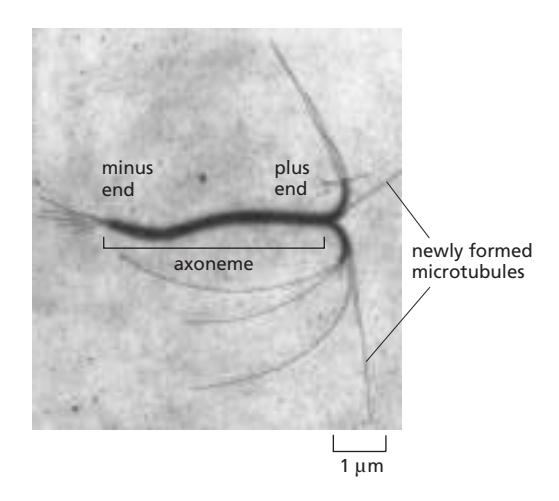
Microtubule은 minus end 쪽(α-tubulin)보다 plus end 쪽(β-tubulin)에서 더 빠른 성장을 한다. GTP-tubulin이 추가되며, GTP 가수분해 후 GDP-tubulin은 불안정해져 탈중합이 일어난다. Dynamic instability: GTP cap이 유지될 때는 성장, cap이 사라지면 catastrophe(급격한 탈중합) → rescue 사이클을 반복한다.
약물 저해제 (→ Chemical Inhibitors of Actin and Microtubules 참조):
| Chemical | Effect | Mechanism | Source |
|---|---|---|---|
| Taxol (paclitaxel) | Stabilizes | Binds along filaments | Yew tree |
| Nocodazole | Depolymerizes | Binds tubulin subunits | Synthetic |
| Colchicine | Depolymerizes | Caps filament ends | Autumn crocus |
- Taxol: microtubule 측면에 결합하여 과안정화 → 방추사 disassembly 방해 → 염색체를 양 극으로 잡아당기지 못하게 함 → 항암 효과 (특히 유방암·난소암)
- Nocodazole: tubulin subunit에 결합하여 중합 억제
- Colchicine: tubulin subunit에 결합하여 필라멘트 말단 capping → 중합 저해
중합 저해제와 탈중합 저해제 모두 mitotic spindle의 올바른 dynamics가 필수적이기 때문에 분열 세포를 우선적으로 사멸시킨다.
Centrosome (MTOC)
→ Centrosome 참조
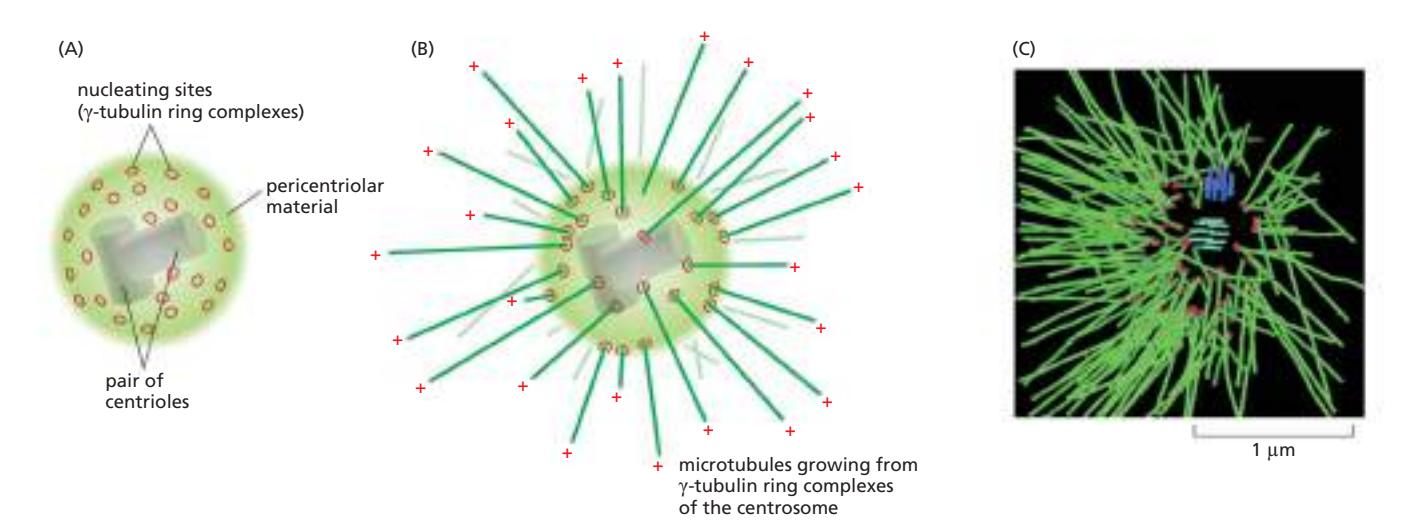 Fig 16–47. Centrosome 구조. (A) 핵 옆에 위치하며, centriole 쌍을 pericentriolar material이 둘러싸는 구조. (B) Microtubule이 부착된 centrosome. Minus end는 centrosome에 묻히고 plus end는 세포질 쪽으로 자유롭게 뻗는다.
Fig 16–47. Centrosome 구조. (A) 핵 옆에 위치하며, centriole 쌍을 pericentriolar material이 둘러싸는 구조. (B) Microtubule이 부착된 centrosome. Minus end는 centrosome에 묻히고 plus end는 세포질 쪽으로 자유롭게 뻗는다.
Centrosome은 동물 세포에서 가장 중요한 **MTOC (microtubule-organizing center)**로, 핵 옆 세포질에 위치한다. Minus end를 고정하고 plus end를 세포 바깥쪽으로 향하게 하여 microtubule을 nucleate하며, 세포 내 organelle 배치의 전체적인 좌표계를 제공한다. Microtubule 합성은 plus end에서 일어나고, minus end에서는 고정만 한다.
γ-Tubulin Ring Complex (γ-TuRC):
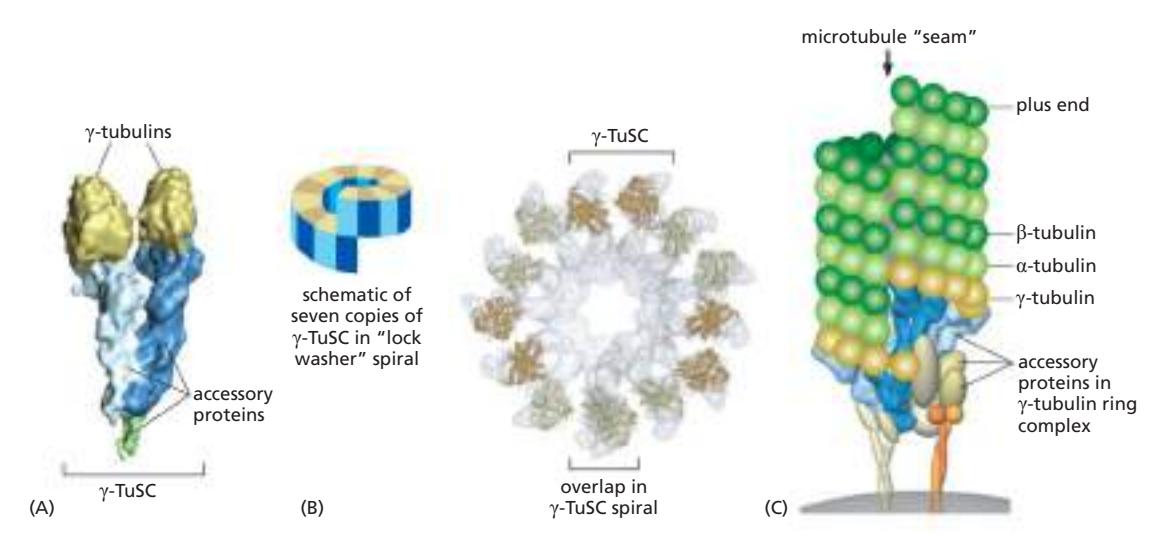 Fig 16–41. γ-TuRC. (B) 7개의 γ-TuSC가 나선형으로 모여 13개의 γ-tubulin이 원형으로 노출. (C) γ-TuRC가 microtubule minus end를 nucleate하는 모델.
Fig 16–41. γ-TuRC. (B) 7개의 γ-TuSC가 나선형으로 모여 13개의 γ-tubulin이 원형으로 노출. (C) γ-TuRC가 microtubule minus end를 nucleate하는 모델.
γ-tubulin 두 개 + 한 쌍의 accessory protein → γ-TuSC (γ-tubulin small complex)7개의 γ-TuSC → 나선형 구조로 13개의 γ-tubulin이 원형으로 노출이것이 microtubule의 13 protofilament와 일치하는 주형을 제공하여 minus end를 nucleate
Pericentriolar Material (PCM): centriole 주변을 감싸는 섬유성 단백질 무정형 망상 구조. γ-TuRC가 여기에 묻혀 microtubule nucleation이 일어난다.
Centrioles
→ Centriole 참조
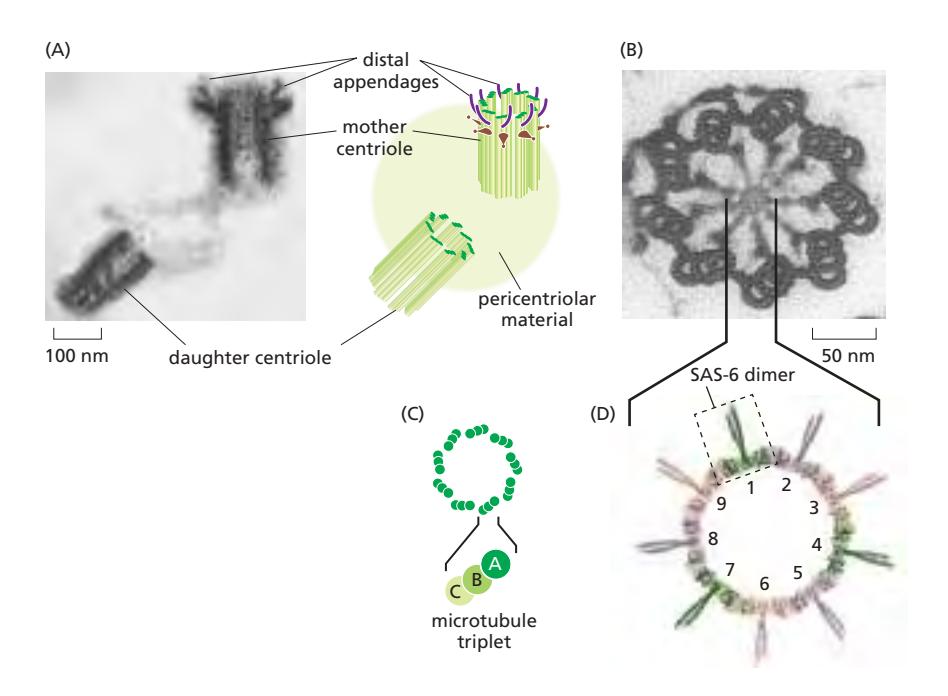 Fig 16–48. (A) Distal appendage를 가진 mother centriole과 인접한 daughter centriole. (B) Protozoan centriole 단면: 9세트의 microtubule triplet. (C) Triplet 구성. (D) SAS-6 단백질이 cartwheel의 9중 대칭 형성.
Fig 16–48. (A) Distal appendage를 가진 mother centriole과 인접한 daughter centriole. (B) Protozoan centriole 단면: 9세트의 microtubule triplet. (C) Triplet 구성. (D) SAS-6 단백질이 cartwheel의 9중 대칭 형성.
- Centrosome 내부에 L자형으로 배치된 한 쌍의 centriole이 있음
- 각 centriole은 9세트의 microtubule triplet이 원통형으로 배열된 구조 (9중 대칭)
- 각 triplet: 하나의 완전한 microtubule (A microtubule) + 두 개의 불완전한 microtubule (B, C microtubule)이 융합
- 중앙 cartwheel의 9중 대칭: SAS-6 단백질이 coiled-coil dimer를 이루어 9개가 자기조립하여 링 형성
| Mother centriole | Daughter centriole | |
|---|---|---|
| 형성 시기 | 이전 세포주기 | 현재 S phase |
| 구조적 특징 | Distal appendage 보유 | Appendage 없음 |
| 기능적 차이 | Primary cilia의 basal body로 기능 | 분열 후 새로운 mother가 됨 |
Centrosome 복제와 세포 분열: S phase에 복제 → mitosis 시 두 centrosome이 분리되어 mitotic spindle 양극 형성
Centrosome의 이중 기능: interphase에서 mother centriole은 세포막 근처로 이동 → primary cilia의 basal body로 기능
- 분열 후 (Interphase): Mother centriole이 세포막 근처로 이동 → primary cilium 형성
- 분열 준비 (S phase): Primary cilium 흡수 → centriole 복제
- 분열기 (Mitosis): 두 centrosome이 spindle 양극 형성
Two Types of Motor Proteins Move Along Microtubules
microtubule-based motor protein의 기능:
- Organelle과 macromolecule 등의 cargo 수송
- Microtubule 간 sliding
- 신경세포, 상피세포 등에서 특이적 microtubule 배열 생성
- Microtubule dynamics 조절 (예: kinesin-13)
Kinesin
→ kinesin 참조
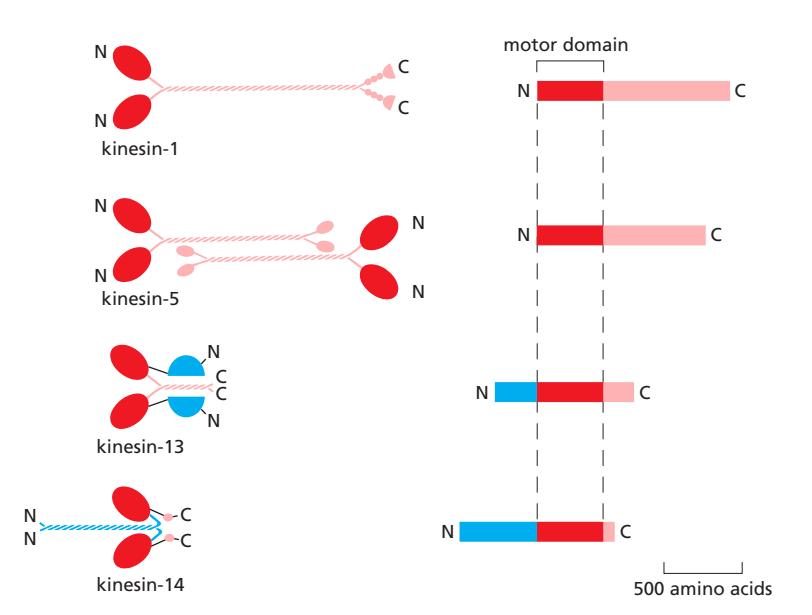 Fig 16–56. Kinesin superfamily. Kinesin-1 (N-terminal motor, plus end), kinesin-5 (tetramer, MT sliding), kinesin-13 (중앙 motor, depolymerization), kinesin-14 (C-terminal motor, minus end).
Fig 16–56. Kinesin superfamily. Kinesin-1 (N-terminal motor, plus end), kinesin-5 (tetramer, MT sliding), kinesin-13 (중앙 motor, depolymerization), kinesin-14 (C-terminal motor, minus end).
Kinesin-1 구조: 두 개의 heavy chain으로 이루어진 이합체. 각 heavy chain은 N-terminal globular head (motor domain) + coiled-coil tail (dimerization) + C-terminal tail (cargo 결합) 구조. Motor domain이 β-tubulin에 결합. Plus end 방향으로 이동.
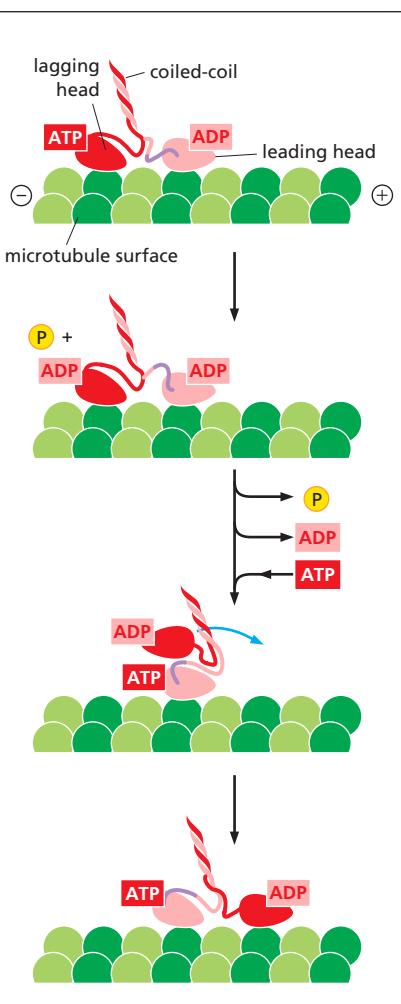 Fig 16–53. Kinesin mechanochemical cycle. “Hand-over-hand” 방식으로 이동. 각 step은 8 nm.
Fig 16–53. Kinesin mechanochemical cycle. “Hand-over-hand” 방식으로 이동. 각 step은 8 nm.
Hand-over-hand stepping 메커니즘:
- 한 head(후방, lagging head)가 ATP와 함께 microtubule에 단단히 결합
- 다른 head(전방, leading head)는 ADP와 함께 느슨하게 결합
- Leading head에 ATP 결합 → neck linker가 앞으로 이동 → 후방 head를 앞으로 당김
- ATP 가수분해 + Pi 방출 후 후방 head가 분리 → 새 위치로 이동
- Processively하게 plus end를 향해 이동
Kinesin Family 다양성:
- N-terminal motor domain → plus end 방향 (대부분의 kinesin)
- C-terminal motor domain → minus end 방향 (kinesin-14)
- 중앙 motor domain → 이동하지 않고 microtubule end에서 depolymerization 촉진 (kinesin-13)
세포 내에서 kinesin은 organelle과 vesicle을 세포 중심에서 periphery로 수송(centrifugal); anterograde axonal transport 담당 (mitochondria, synaptic vesicle 전구체 등).
Dynein
→ dynein 참조
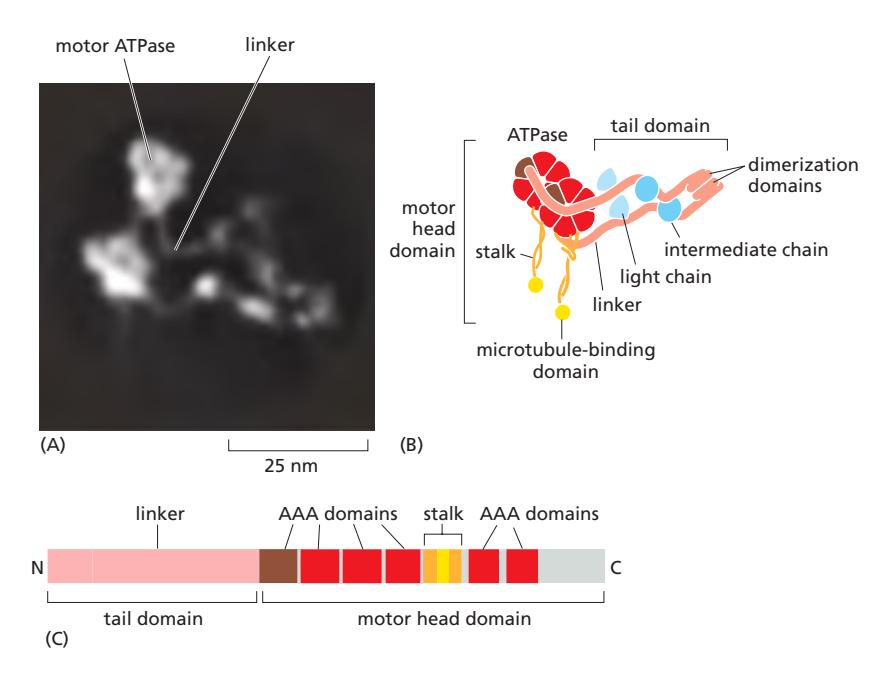 Fig 16–58. Cytoplasmic dynein. (A) cryoEM 재구성. (B) 두 heavy chain: motor head, stalk, tail domain 구성.
Fig 16–58. Cytoplasmic dynein. (A) cryoEM 재구성. (B) 두 heavy chain: motor head, stalk, tail domain 구성.
Dynein은 minus end 방향으로 이동하는 motor protein 계열이다. Dynein의 두 major branch: cytoplasmic dynein과 ciliary (axonemal) dynein. Dynein의 motor domain은 myosin과 kinesin의 것보다 크다.
Cytoplasmic dynein 1: 두 개의 heavy chain으로 이루어진 homodimer. 거의 모든 진핵세포에 단일 유전자로 인코딩. Organelle과 mRNA 수송, mitosis에서 spindle 구성, 세포 이동 시 centrosome과 핵의 위치 결정에 관여. Cargo에 결합하기 위해 dynactin 복합체와 adaptor protein이 필요.
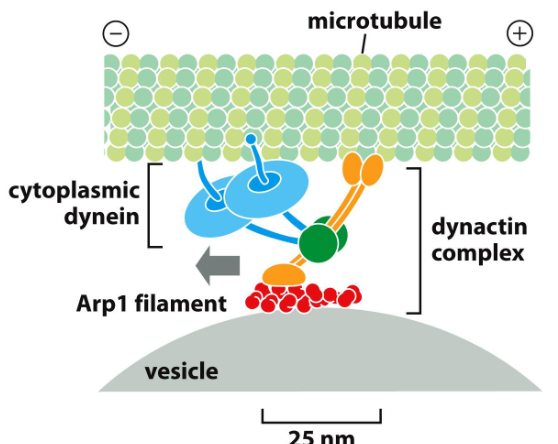 Dynein은 vesicular organelle 운반을 위해 다수의 accessory protein을 필요로 함.
Dynein은 vesicular organelle 운반을 위해 다수의 accessory protein을 필요로 함.
Dynactin: dynein과 microtubule, actin-like filament (Arp1으로 구성됨)에 각각 결합하는 요소들로 구성됨. Membrane-enclosed organelle에 dynein이 결합하는 것을 돕는다.
Microtubule Organization in a Neuron
→ Microtubule organization in a neuron 참조
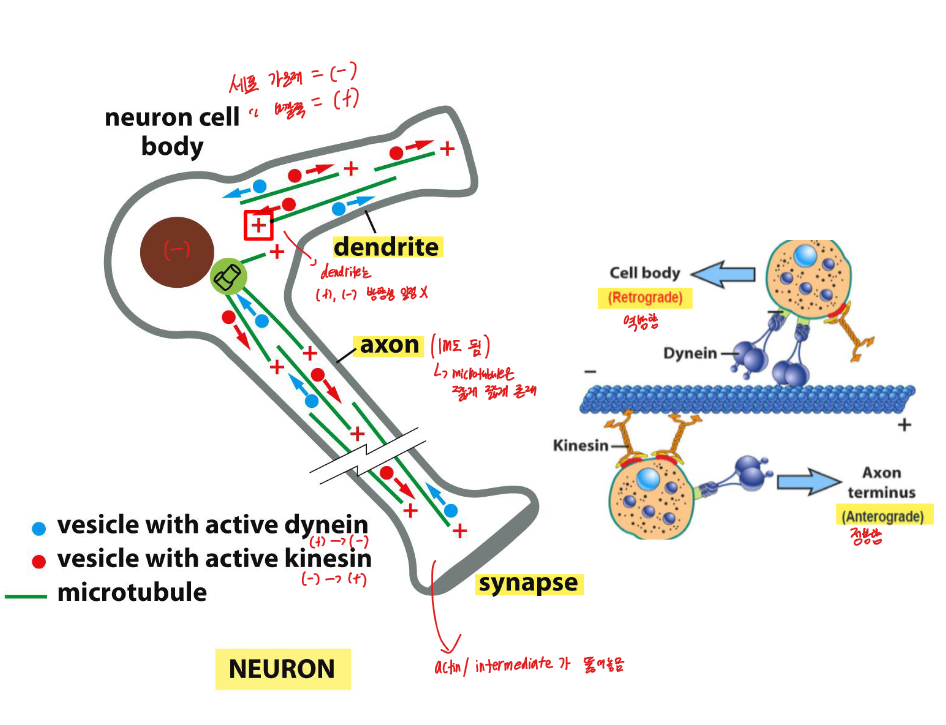 Fig 16–62. 뉴런에서 microtubule 조직. 축삭: 모든 plus end가 말단 쪽. 수상돌기: 혼재 극성.
Fig 16–62. 뉴런에서 microtubule 조직. 축삭: 모든 plus end가 말단 쪽. 수상돌기: 혼재 극성.
Axon (축삭):
- 모든 microtubule이 같은 극성 공유 — plus end가 축삭 말단(axon terminus) 쪽을 향함
- 하나의 microtubule이 축삭 전체 길이를 뻗지 않으며, 짧게 겹치는 평행 microtubule 세그먼트들이 fast axonal transport의 track 제공
- Anterograde axonal transport (세포체 → 말단): plus end 방향 → kinesin이 담당 (mitochondria, synaptic vesicle 전구체 등)
- Retrograde axonal transport (말단 → 세포체): minus end 방향 → cytoplasmic dynein이 담당
Dendrite (수상돌기):
- Microtubule 극성이 혼재 — plus end가 바깥과 안쪽을 향하는 것이 섞임
- Vesicle이 kinesin과 dynein 모두에 결합 가능, 어떤 motor가 활성화되느냐에 따라 양방향 이동
알츠하이머 병리: 세포 내 microtubule을 안정화하는 Tau 단백질이 과인산화되면 미세소관이 붕괴하여 synapse까지 수송이 어려워지면서 neuronal malfunction이 발생한다.
Flagellum and Cilium (9+2 Structure)
→ 320_Flagellum or cilium_MOC 참조
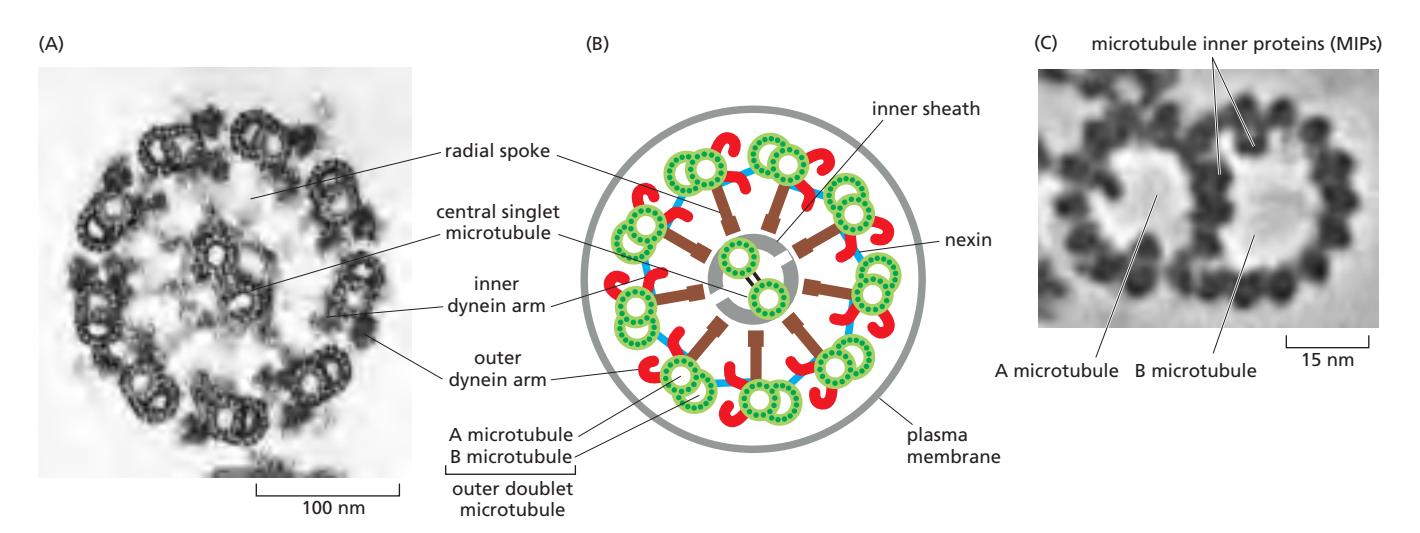 Fig 16–58. 편모/섬모 구조. (A) Chlamydomonas 편모 단면: “9+2” microtubule 배열. (B) 편모/섬모 부분별 구조 모식도. (C) 외부 microtubule doublet 고해상도 tomography.
Fig 16–58. 편모/섬모 구조. (A) Chlamydomonas 편모 단면: “9+2” microtubule 배열. (B) 편모/섬모 부분별 구조 모식도. (C) 외부 microtubule doublet 고해상도 tomography.
Flagella와 cilia는 hairlike 세포 돌출물로, 핵심에 axoneme이라 불리는 microtubule 기반 구조를 공유한다. 원생동물에서 인간까지 거의 모든 진핵 편모/섬모에서 동일한 배열이 보존된다.
Axoneme 구조 (9+2 배열)
- 9개의 doublet microtubule이 원형으로 배열되고, 중앙에 **단일 microtubule 쌍(central pair)**이 위치
- Doublet: 완전한 A microtubule + 부분적인 B microtubule이 공통 벽을 공유하며 융합된 구조
- Microtubule은 axoneme 전체 길이(10–200 μm)를 따라 뻗으며, 규칙적 간격으로 accessory protein이 cross-link
Accessory Proteins
| 구조 | 위치 | 기능 |
|---|---|---|
| Inner dynein arm | A microtubule → 인접 B microtubule | Bending 운동 생성 (ciliary dynein) |
| Outer dynein arm | A microtubule → 인접 B microtubule | Bending 운동 생성 (ciliary dynein) |
| Nexin (conexin) | Doublet 간 연결 | 9개 doublet 간 간격 유지; sliding을 억제하여 bending 가능하게 함 |
| Radial spoke | A microtubule → central pair | Doublet과 중앙 쌍을 연결; |
Ciliary Dynein (Axonemal Dynein)
→ Ciliary dynein 참조
 Fig 16–59. Axonemal dynein. Dynein arm이 한 doublet의 A microtubule과 인접 doublet의 B microtubule을 규칙적 간격으로 연결.
Fig 16–59. Axonemal dynein. Dynein arm이 한 doublet의 A microtubule과 인접 doublet의 B microtubule을 규칙적 간격으로 연결.
Ciliary dynein (섬모 dynein)은 axoneme 내 인접 microtubule doublet을 연결하는 minus end 방향 motor이다.
Dimeric 또는 trimeric 구조; 1~3개의 motor-containing heavy chainTail: A microtubule에 단단히 결합Globular head: stalk을 통해 인접 B microtubule에 ATP-의존적으로 결합- Ciliary dynein 한쪽은 A tubulin, 나머지 한쪽은 반대편 doublet의 B tubulin에 결합
Axoneme Bending
→ axoneme bending 참조
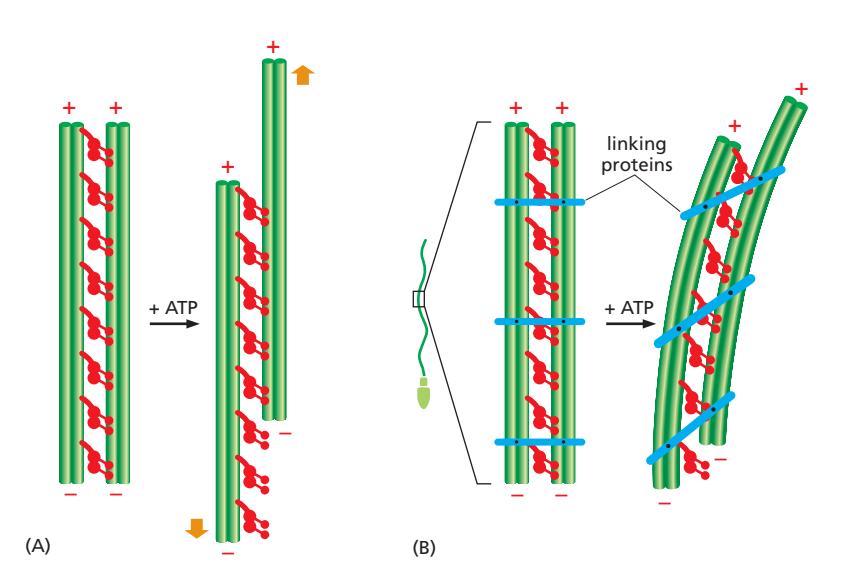 Fig 16–65. Axoneme bending. (A) Trypsin 처리로 cross-link를 끊으면 ATP 추가 시 doublet 간 sliding 발생. (B) Intact axoneme에서는 flexible protein link(conexin)들이 sliding을 막아 dynein 힘이 bending으로 전환.
Fig 16–65. Axoneme bending. (A) Trypsin 처리로 cross-link를 끊으면 ATP 추가 시 doublet 간 sliding 발생. (B) Intact axoneme에서는 flexible protein link(conexin)들이 sliding을 막아 dynein 힘이 bending으로 전환.
Dynein head가 ATP를 가수분해하면 B microtubule의 minus end 방향으로 이동하려 하여 인접 doublet 사이에 sliding force를 발생시킨다. 그러나 intact axoneme에서는 doublet 사이를 연결하는 **flexible protein link(conexin)**들이 이 sliding을 억제한다. Sliding이 억제된 상태에서 dynein 힘은 bending 운동으로 전환된다.
- 비대칭적 dynein 활성화가 wave-like 운동을 만들어냄
- 실험적 확인: axoneme을 proteolytic enzyme (trypsin)으로 처리하면 doublet 간 연결 단백질이 끊어짐 → 이 상태에서 ATP 추가 시 실제 sliding 발생. 반면 intact axoneme에서 ATP 추가 시 bending 발생
Primary Cilia
→ primary cilia 참조
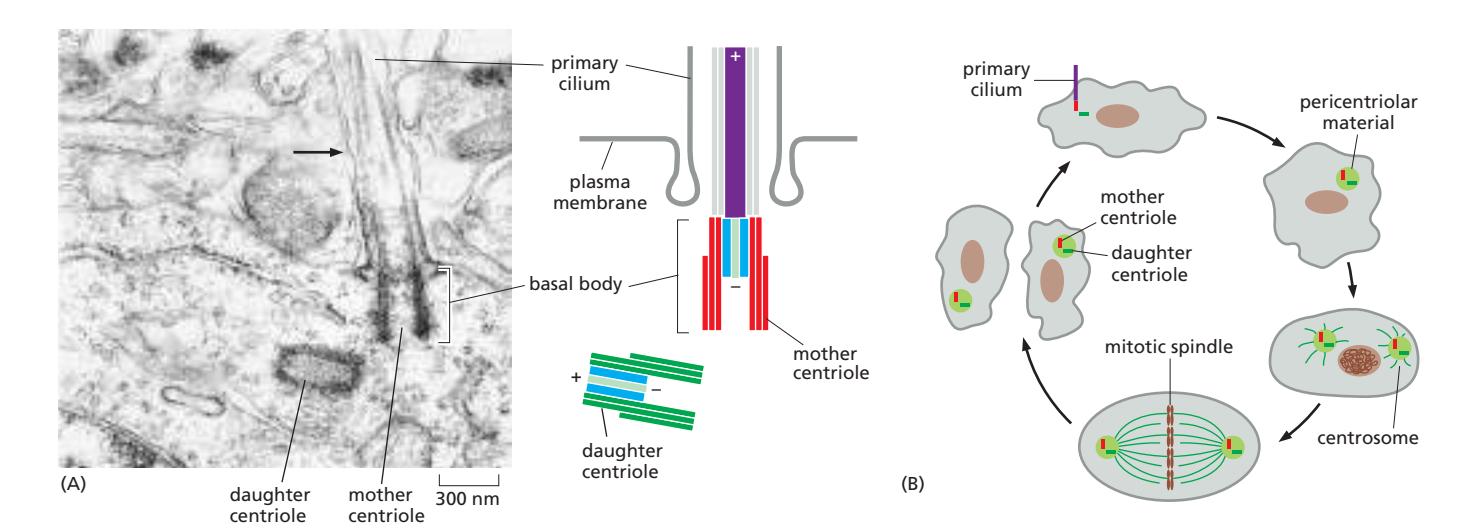 Fig 16–66. Primary cilia. (A) 마우스 뉴런의 primary cilium basal body. (B) Centriole이 basal body와 centrosome core로 번갈아 기능하는 주기.
Fig 16–66. Primary cilia. (A) 마우스 뉴런의 primary cilium basal body. (B) Centriole이 basal body와 centrosome core로 번갈아 기능하는 주기.
Primary cilium은 거의 모든 세포 유형의 표면에서 발견되는 짧고 비운동성인 감각 organelle이다. secondary cilia와 달리 9+0 배열(중앙 pair 없음)을 가진다.
- 형성: interphase 동안 basal body (단일 centriole이 핵심인 구조)에서 plasma membrane 근처에 형성
- Centriole은 세포 분열 시 centrosome의 핵심으로, interphase에서는 basal body로 번갈아 기능
- 세포가 분열 주기에 진입하면 primary cilium이 **흡수(resorption)**되어 centriole이 centrosome으로 기능
Intermediate Filaments
개요
Intermediate filaments는 직경 약 10 nm의 로프 같은(ropelike) 섬유이며, intermediate filament 단백질이라는 크고 이질적인 단백질 패밀리로 만들어진다. 한 유형은 내핵막 바로 아래의 nuclear lamina를 형성하고, 다른 유형들은 세포질 전반에 걸쳐 뻗어 세포에 기계적 강도를 제공한다. 상피 조직에서는 세포질을 가로질러 세포-세포 junction에서 다른 junction으로 연결되어 상피 전체를 강화한다.
Construction of Intermediate Filament
→ construction of intermediate filament 참조
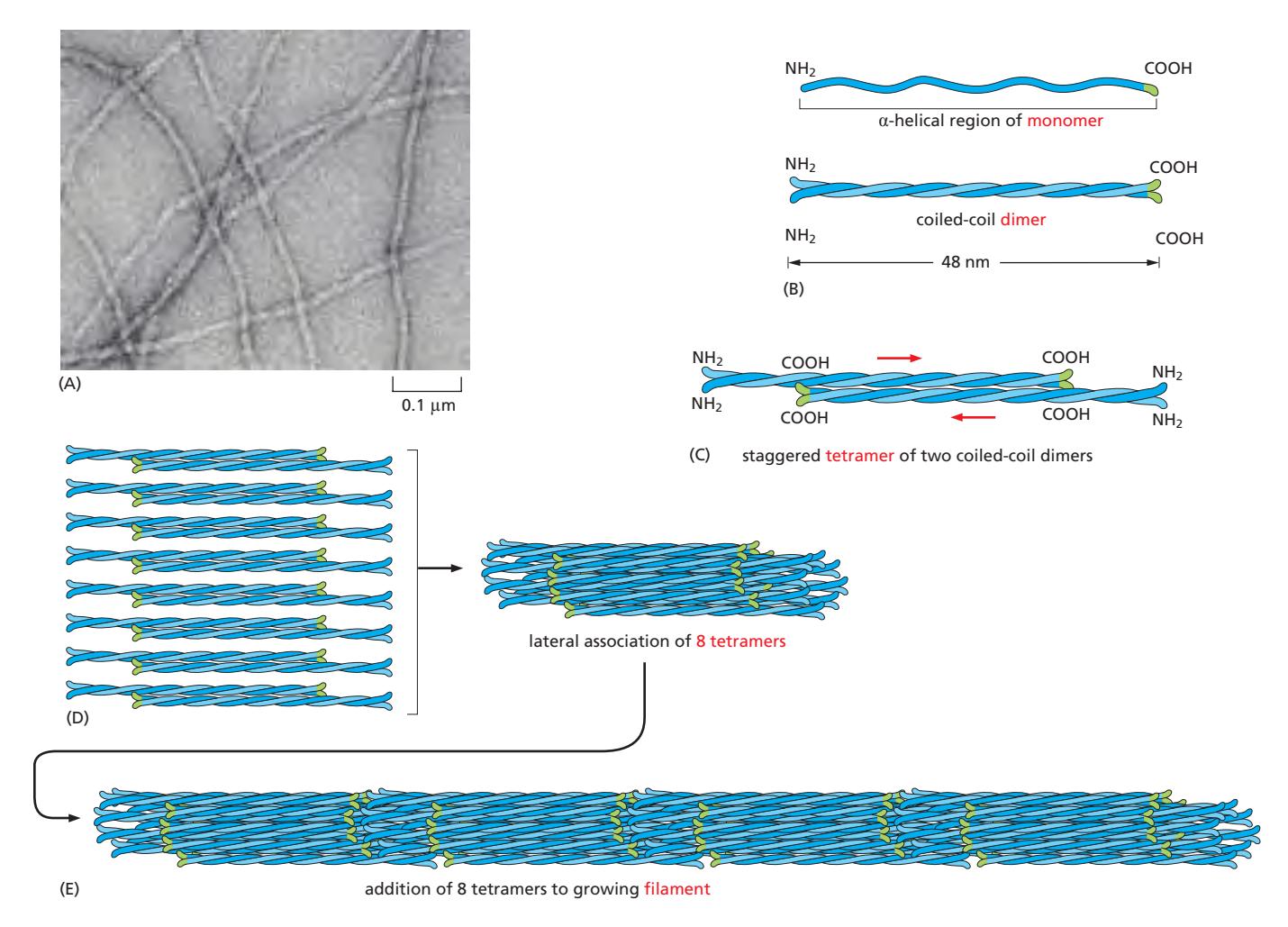 Fig 16–67. Intermediate filament 조립 모델. (B) Monomer. (C) Coiled-coil dimer. (D) Antiparallel staggered tetramer. (E) 10 nm 필라멘트.
Fig 16–67. Intermediate filament 조립 모델. (B) Monomer. (C) Coiled-coil dimer. (D) Antiparallel staggered tetramer. (E) 10 nm 필라멘트.
조립 단계:
- Monomer(α-helix) → Coiled-coil dimer: 두 monomer가 평행하게 정렬되어 central rod domain끼리 coiled-coil 형성
- Dimer → Antiparallel staggered tetramer: 두 dimer가 역방향(antiparallel)으로 엇갈리게 결합하여 4-chain tetramer 형성 (soluble subunit)
- Tetramer → 10 nm 필라멘트: Tetramer들이 측면으로 packing → 8개의 parallel protofilament → 최종 필라멘트 단면에 16개의 dimer (32개의 coiled-coil)
중요한 특성:
- Intermediate filament subunit은 ATP나 GTP 결합 부위가 없음 (actin, microtubule과 구분)
- Tetramer subunit은 두 dimer가 반대 방향으로 구성 → 양 끝이 동일 → assembled intermediate filament는 극성(polarity)이 없음
- 쉽게 구부러지지만(persistence length < 1 μm) 끊기가 매우 어렵고, 원래 길이의 3배 이상 늘어날 수 있음 (rope-like 기계적 특성)
- Assembly/disassembly는 단백질 인산화가 조절; mitosis 시 nuclear lamin 분해가 대표적 예
Types of Intermediate Filament
→ types of intermediate filament 참조
척추동물의 intermediate filament 단백질은 세포 유형과 위치에 따라 분류된다 (Table 16–2):
| 유형 | 구성 단백질 | 발현 위치 |
|---|---|---|
| Nuclear | Lamin A, B, C | 핵막 내면의 nuclear lamina |
| Epithelial | Type I keratin (acidic) | 상피세포 및 그 파생 조직 (모발, 손발톱 등) |
| Type II keratin (neutral/basic) | 상피세포 및 그 파생 조직 | |
| Axonal | NF-L, NF-M, NF-H | 뉴런의 축삭 |
Nuclear lamin: 가장 오래된 IF 단백질. 내핵막을 지지하는 핵막하 망상 구조(nuclear lamina) 형성. 유사분열 시 인산화에 의해 해체 → 분열 후 재조립.
Keratin 필라멘트: IF 중 가장 다양한 계열 (→ Keratin filaments 참조).
Neurofilament: 뉴런 축삭에 고농도 존재. NF-L, NF-M, NF-H가 공조립하여 heteropolymer 형성. NF-H와 NF-M의 긴 C-terminal tail이 인접 필라멘트를 cross-link하여 정렬된 배열 형성. 축삭 직경은 neurofilament 유전자 발현 수준과 직접 연관 → 신경전달속도에 영향.
 (A) 뉴런 축삭의 neurofilament — NF-H C-terminal extension에 의한 cross-bridge. (B) 신경교세포의 glial filament. (C) 축삭 횡단면 — neurofilament가 microtubule보다 훨씬 많이 분포.
(A) 뉴런 축삭의 neurofilament — NF-H C-terminal extension에 의한 cross-bridge. (B) 신경교세포의 glial filament. (C) 축삭 횡단면 — neurofilament가 microtubule보다 훨씬 많이 분포.
Keratin Filaments
→ Keratin filaments 참조
Keratin은 intermediate filament 중 가장 다양한 계열로, 인간 유전체에 54종의 distinct keratin 존재. 상피세포에서는 약 20종, 모발과 손발톱에는 약 10종이 특이적 발현.
구조와 조립: Type I (acidic) keratin과 Type II (neutral/basic) keratin이 1:1 비율로 결합한 heterodimer로 조립. 이황화 결합(disulfide bond)으로 cross-linked된 keratin 네트워크는 세포가 죽은 후에도 유지 → 피부 각질층, 모발, 손발톱, 비늘 같은 단단한 외피 구조 형성.
이 intermediate filament는 세포 내에서 만들어지며, 특정 세포들은 이들을 세포 내에 쌓아두어 결국 세포가 keratin 덩어리가 되고 각질화된다.
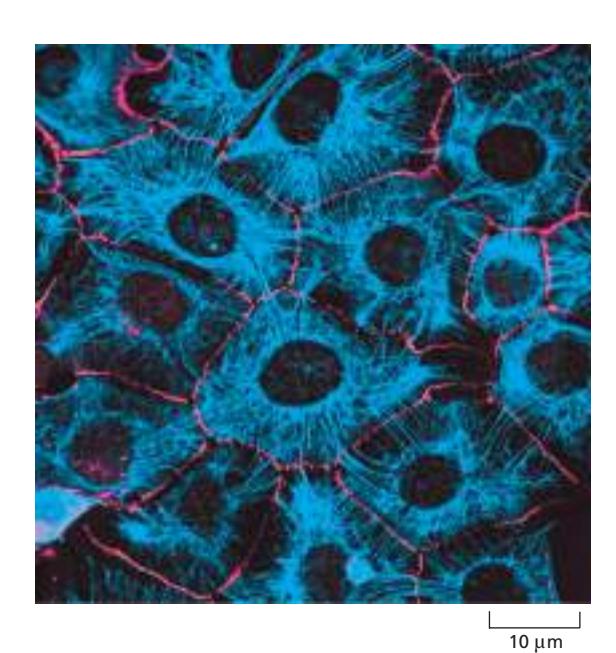 배양된 상피세포 시트에서 keratin 필라멘트 네트워크. 붉은색이 세포막, 푸른색이 keratin filament. Desmosome을 통해 인접 세포와 간접적으로 연결.
배양된 상피세포 시트에서 keratin 필라멘트 네트워크. 붉은색이 세포막, 푸른색이 keratin filament. Desmosome을 통해 인접 세포와 간접적으로 연결.
조직 내 연결: Keratin 필라멘트는 두 곳의 cell junction에 고정되어 상피 조직 전체에 기계적 강도를 전달.
- Desmosome: 세포-세포 접촉부에서 인접 세포의 keratin 네트워크 연결
- Hemidesmosome: 세포-기질 접촉부에서 keratin 네트워크를 basal lamina에 연결
Epidermolysis Bullosa Simplex (단순수포성표피박리증):
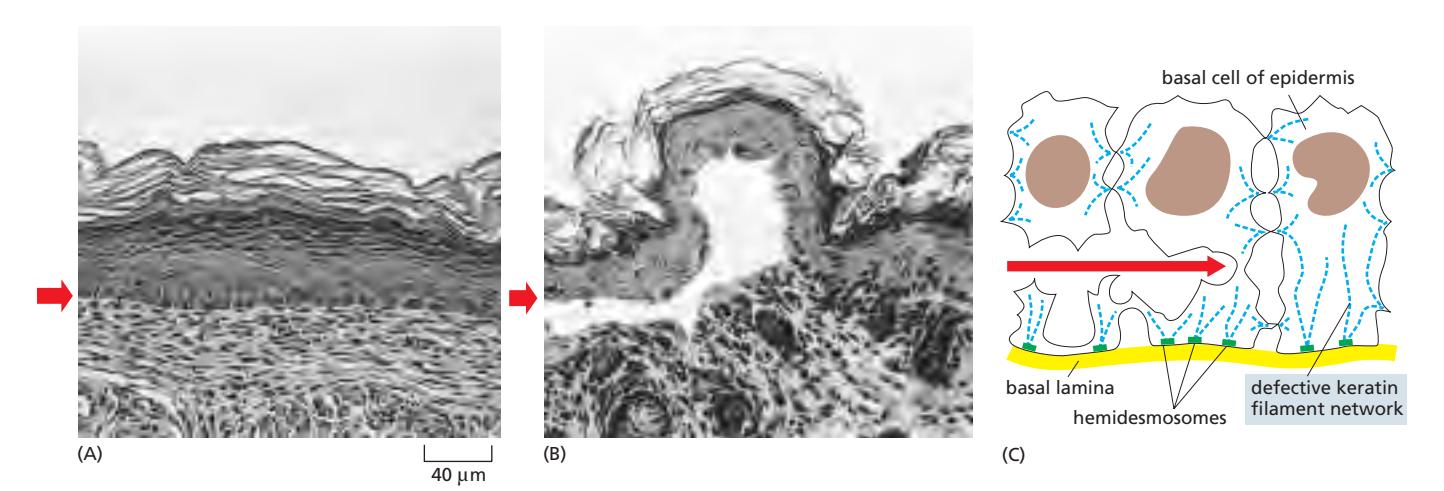 (A) 정상 피부 단면. (B) Dominant negative mutation keratin을 발현하는 형질전환 마우스의 피부 — 표피 기저층 세포가 파열되어 수포 형성(빨간 화살표). Gene의 dual copy 중 하나에만 변이가 발생해도 문제가 생김. (C) 기저층 세포 모식도 — 세포는 핵과 hemidesmosome 사이에서 파열.
(A) 정상 피부 단면. (B) Dominant negative mutation keratin을 발현하는 형질전환 마우스의 피부 — 표피 기저층 세포가 파열되어 수포 형성(빨간 화살표). Gene의 dual copy 중 하나에만 변이가 발생해도 문제가 생김. (C) 기저층 세포 모식도 — 세포는 핵과 hemidesmosome 사이에서 파열.
- 표피 기저세포층(basal cell layer)에서 결함 있는 keratin이 발현될 때 발생
- 아주 약한 기계적 자극에도 피부가 수포를 형성하는 것이 특징
- 결함 있는 단백질이 정상 keratin과 공조립되어 필라멘트 네트워크 전반을 교란 (dominant negative 효과)
- Keratin의 중심 rod domain 말단 부위 돌연변이가 이 질환을 일으키는 경우가 많아, 해당 부위가 올바른 필라멘트 조립에 특히 중요함을 보여줌
- 유사 기전으로 구강 점막, 식도 점막, 각막 등 특정 조직에서도 수포 형성 가능
Plectin
→ Plectin 참조
 Plectin(녹색)이 intermediate filament(파랑)과 microtubule(빨강) 사이를 cross-link하는 전자현미경 이미지. 노란 점은 anti-plectin 항체에 연결된 금 입자. Actin filament는 Plectin을 잘 보기 위해 제거되었다.
Plectin(녹색)이 intermediate filament(파랑)과 microtubule(빨강) 사이를 cross-link하는 전자현미경 이미지. 노란 점은 anti-plectin 항체에 연결된 금 입자. Actin filament는 Plectin을 잘 보기 위해 제거되었다.
Plectin은 plakin 계열 단백질로, 대형 modular 단백질이다. 세포골격 필라멘트들을 서로, 그리고 junction complex에 연결하는 여러 domain을 가지며 세포골격 통합의 대표적 예시이다.
연결 대상:
- Intermediate filament 간 bundling
- Intermediate filament ↔ microtubule
- Intermediate filament ↔ actin filament bundle
- Intermediate filament ↔ myosin II 필라멘트
- Intermediate filament bundle ↔ plasma membrane의 adhesive structure (desmosome, hemidesmosome 등)
- SUN-KASH protein complexes와 연결 → intermediate filament를 핵 내부와 간접적으로 연결하는 다리 역할
Plectin의 cross-linking 작용은 세포가 기계적 스트레스를 견디는 데 필수적이다 (Plectin 유전자 없는 마우스: 수포 피부, 비정상적 골격근·심근, 수일 내 사망).
SUN-KASH Protein Complexes (LINC Complex)
→ SUN-KASH protein complexes 참조
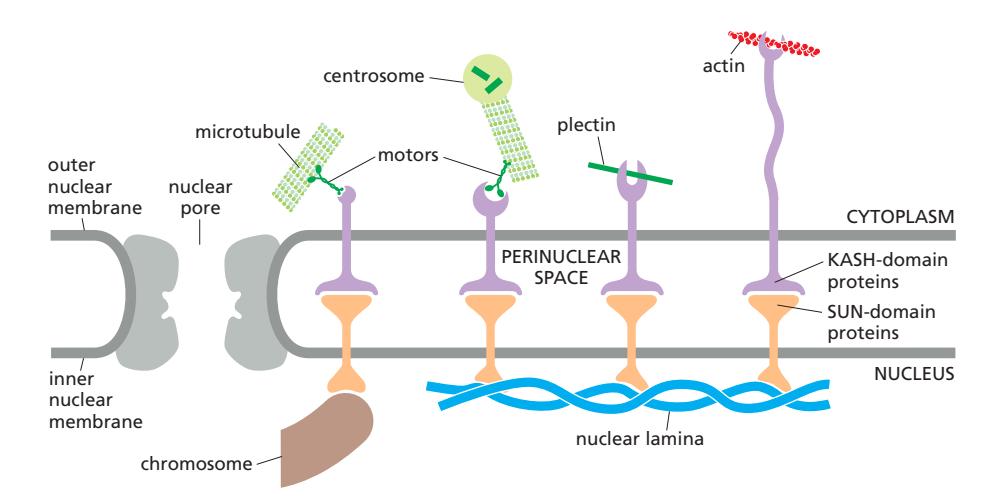 SUN-KASH 복합체(LINC complex)가 핵막을 통해 세포질 세포골격과 핵 내부를 연결. SUN(주황)은 내핵막, KASH(보라)는 외핵막에 위치. 두 단백질의 결합 domain이 핵막 내강(lumen)에서 맞닿는다.
SUN-KASH 복합체(LINC complex)가 핵막을 통해 세포질 세포골격과 핵 내부를 연결. SUN(주황)은 내핵막, KASH(보라)는 외핵막에 위치. 두 단백질의 결합 domain이 핵막 내강(lumen)에서 맞닿는다.
LINC complex (Linker of Nucleoskeleton and Cytoskeleton)는 핵막을 가로질러 핵 내부와 세포질 세포골격을 기계적으로 연결하는 단백질 복합체이다.
구조:
| 단백질 | 위치 | 결합 상대 |
|---|---|---|
| SUN proteins | 내핵막 | 핵질 쪽: nuclear lamina 또는 염색체; 핵막 내강: KASH domain과 결합 (SUN-KASH 사이는 hydrophobic) |
| KASH proteins | 외핵막 | 핵막 내강: SUN domain과 결합; 세포질 쪽: actin filament (직접), microtubule (motor protein 통해), intermediate filament (plectin 통해) |
기능:
- 감수분열 시 염색체 이동 — 핵 내부의 telomere가 세포질 세포골격에 연결되어 이동 주도
- 핵 및 중심체 위치 결정 — 세포 이동, 신경세포 이동 등에서 중요
- 핵 이동 (nuclear migration) — 특히 발생 과정에서 중요
- 전체적인 세포골격 조직화
- 신호 전달 기능
SUN-KASH 연결이 없으면 핵이 세포골격으로부터 기계적으로 분리된다.